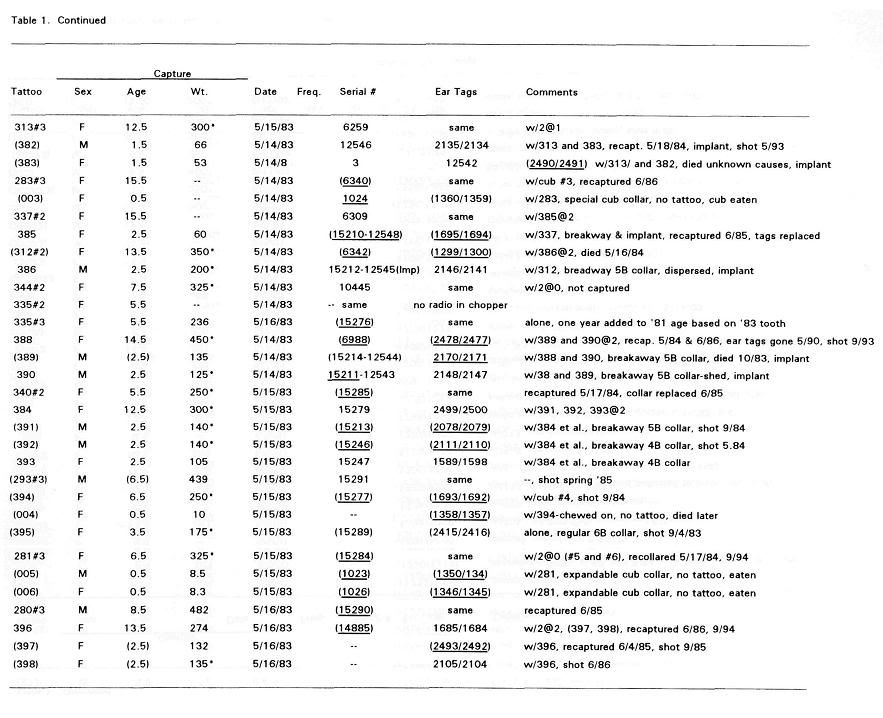
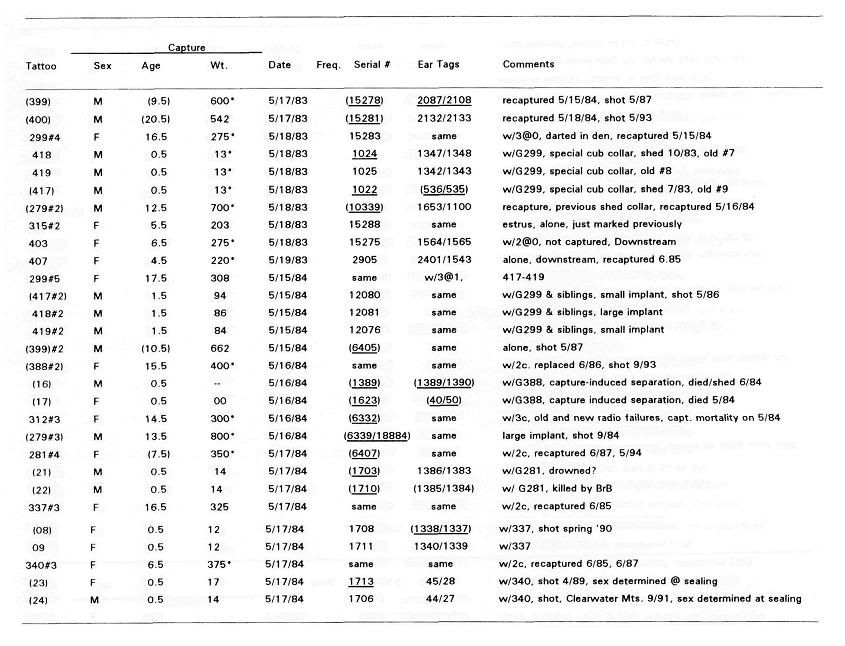
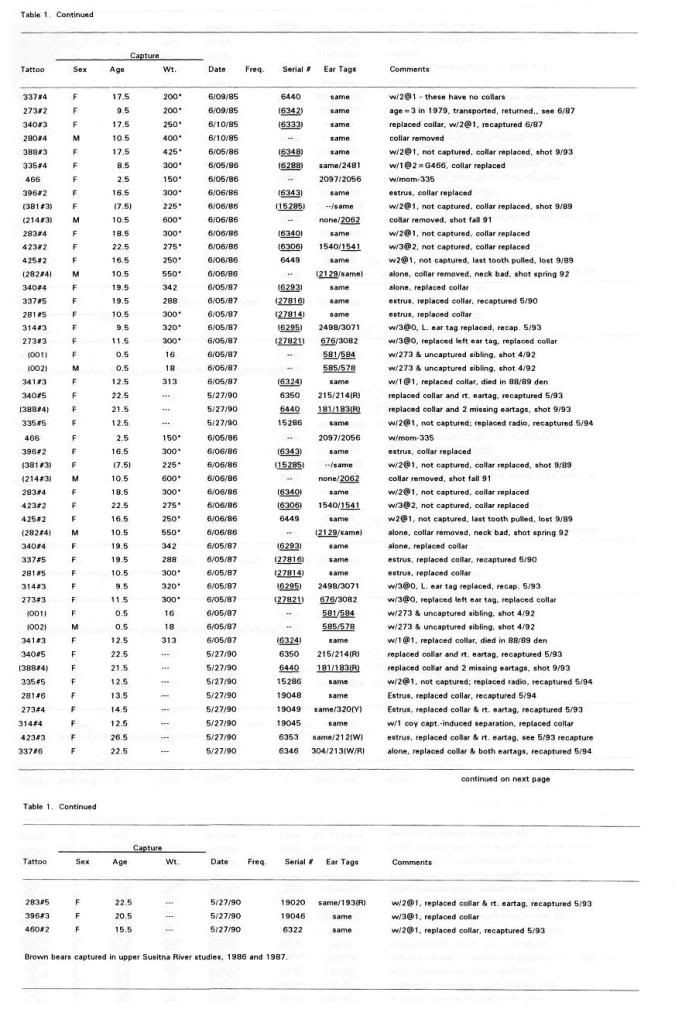
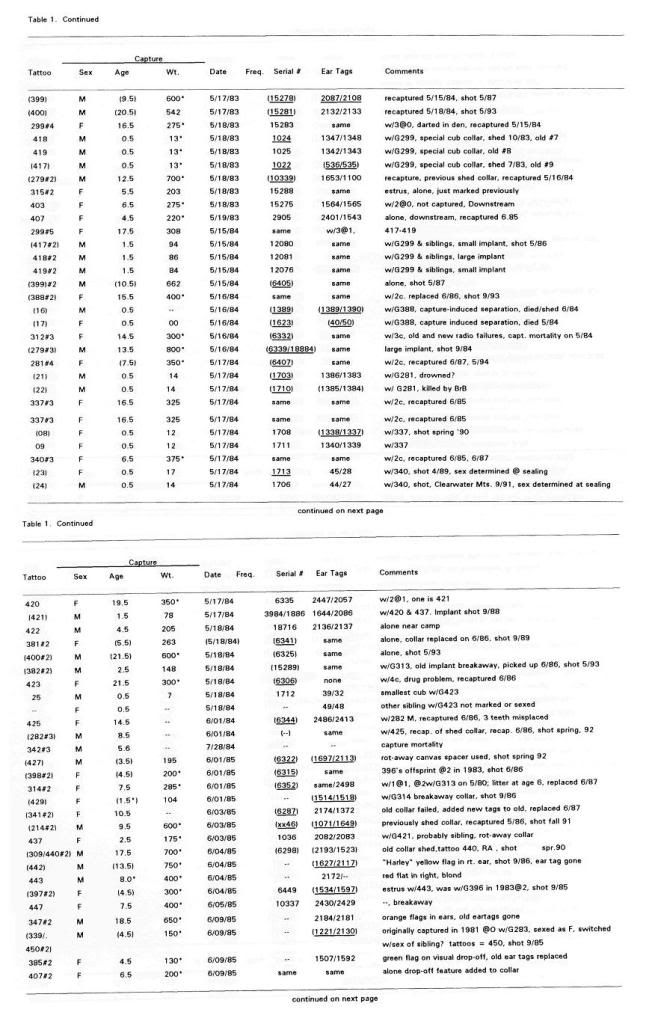
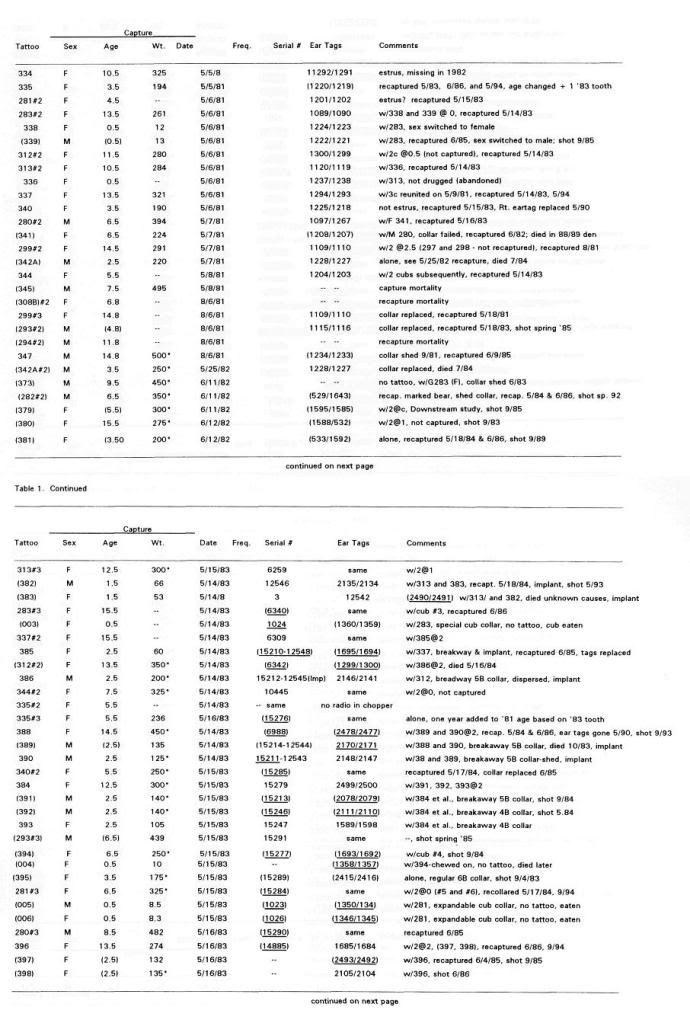
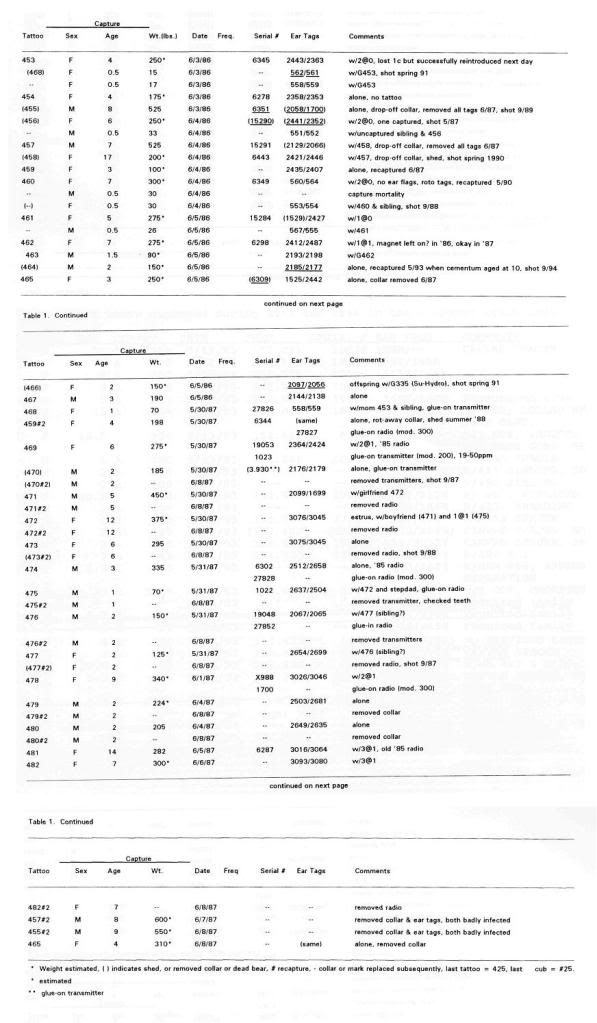
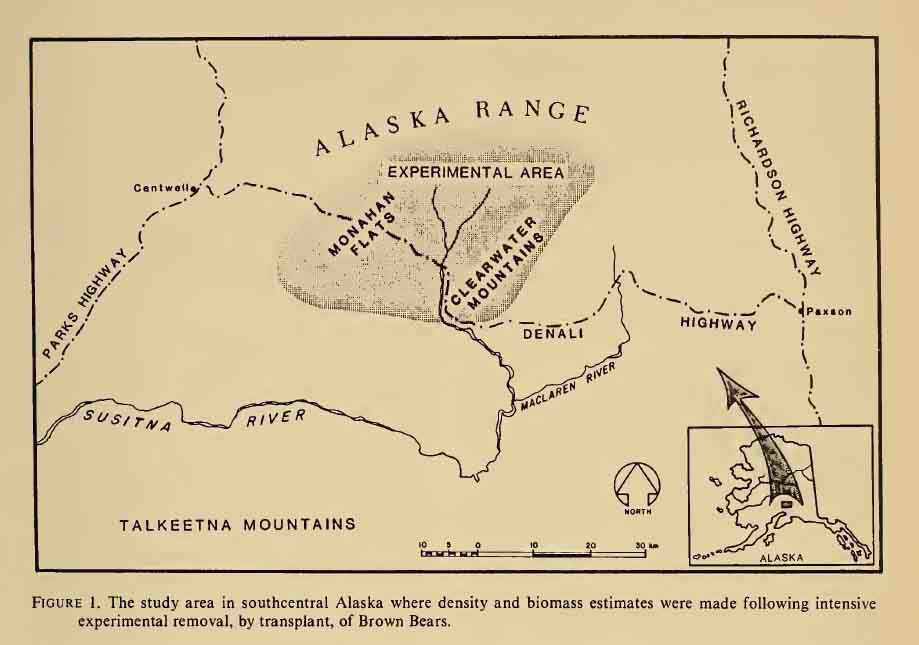
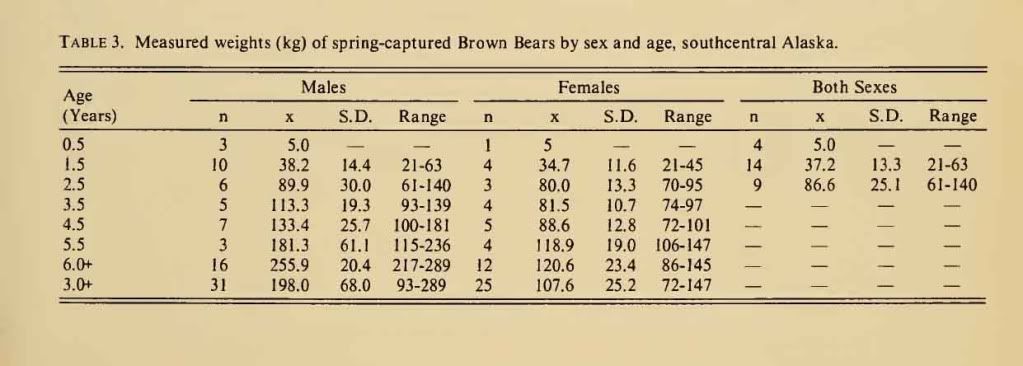
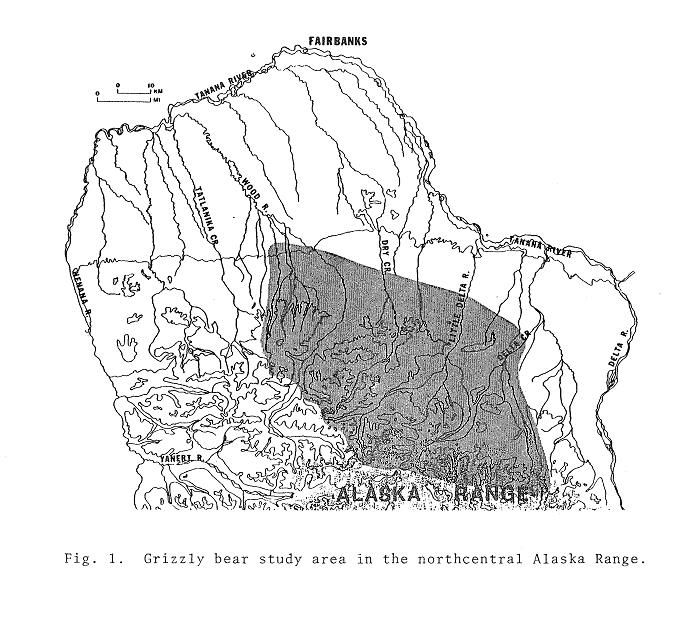
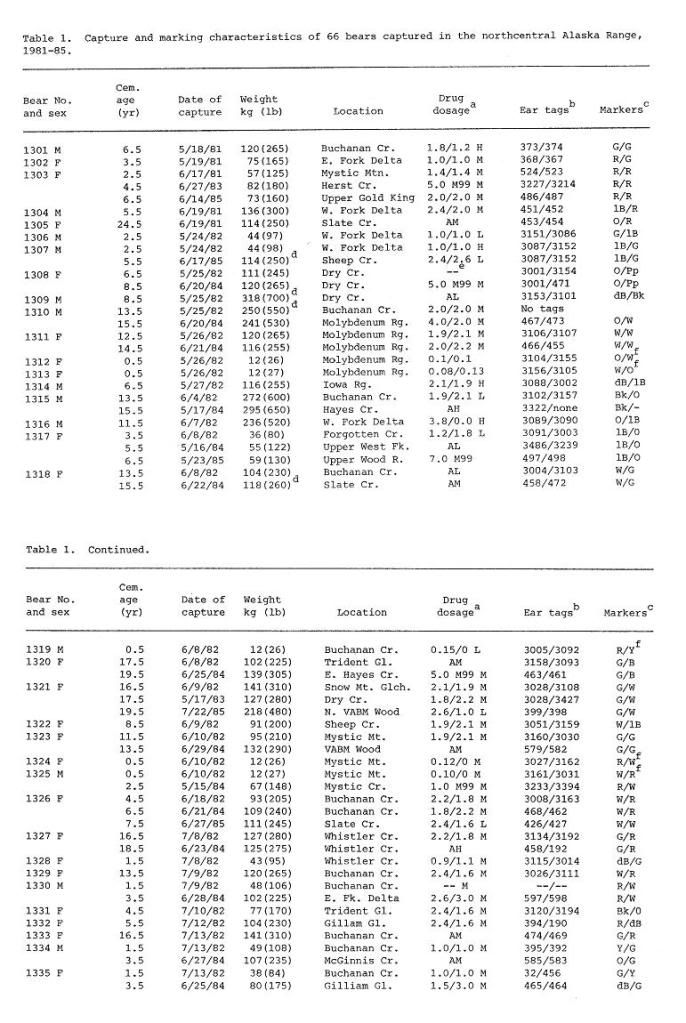
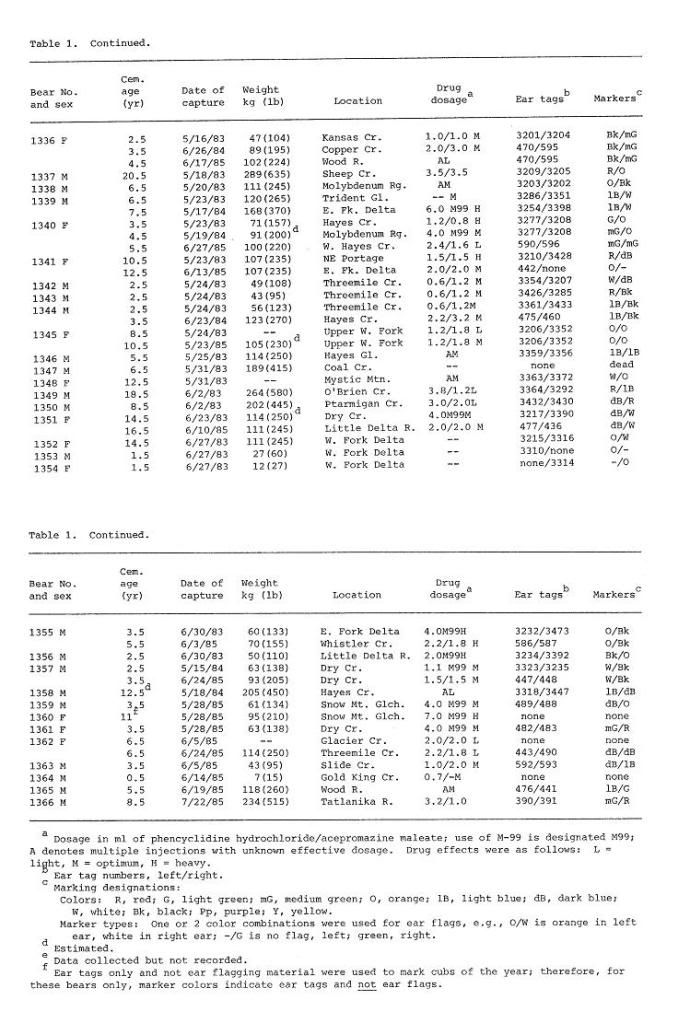
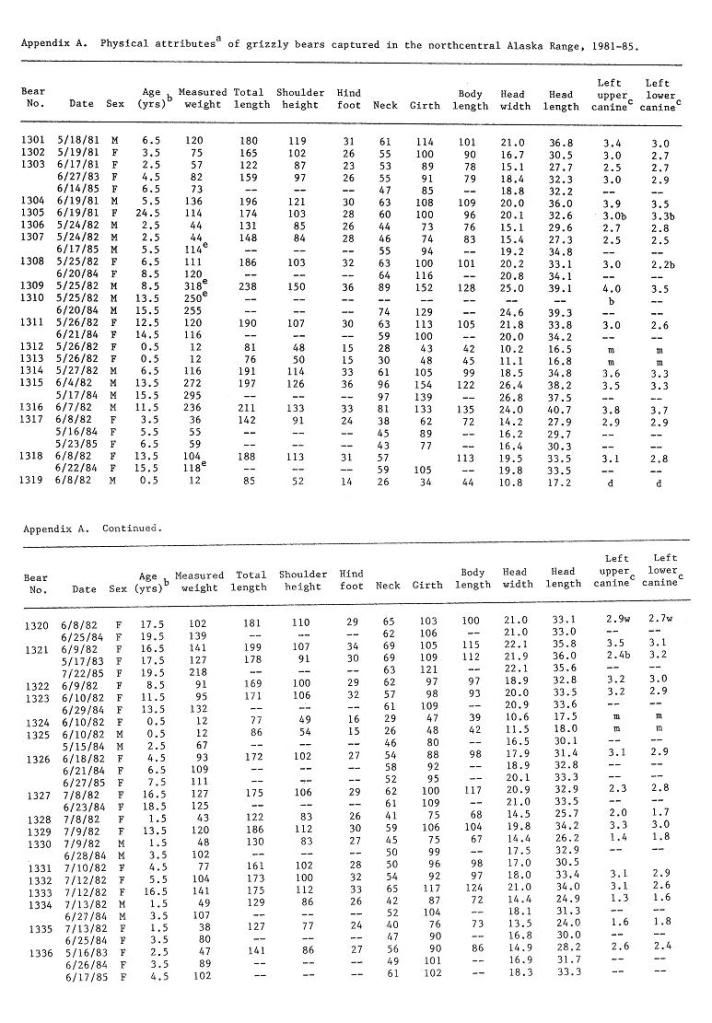
Post by grrraaahhh on Apr 28, 2011 6:54:37 GMT -9
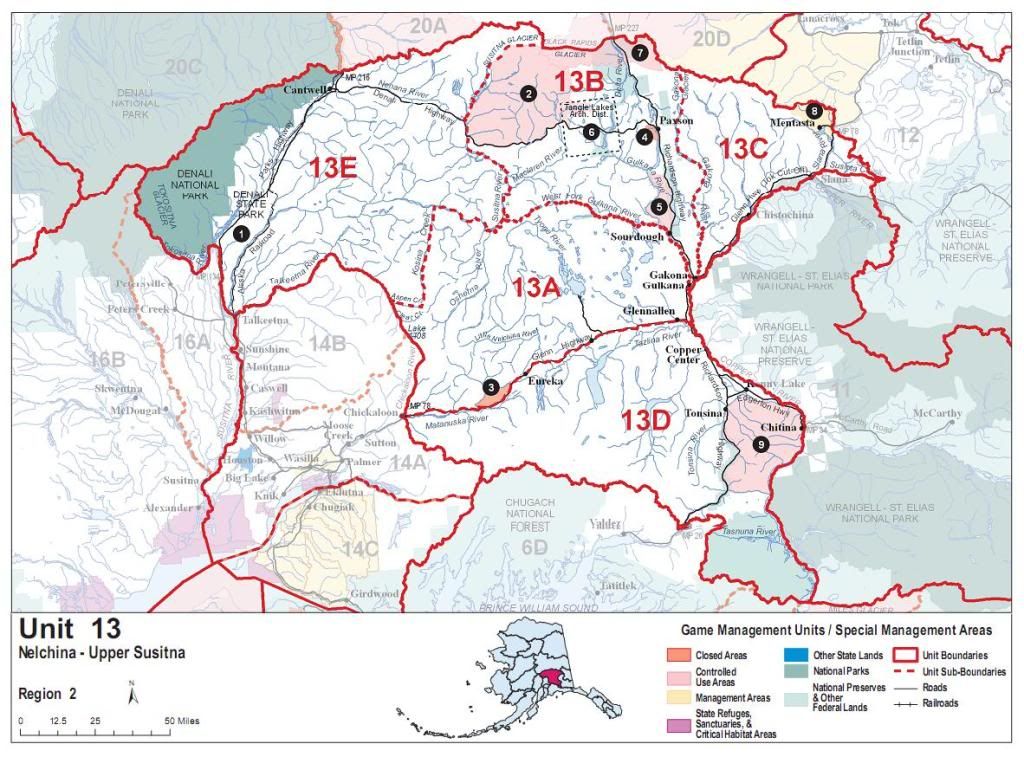
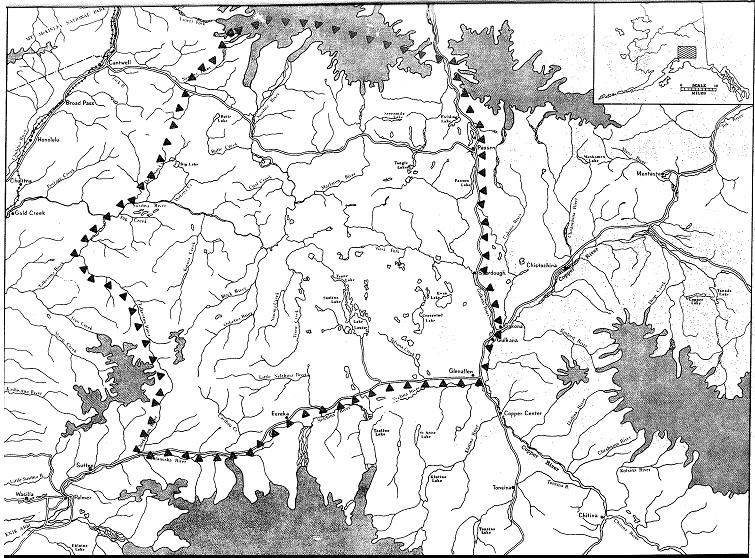
Southwest Yukon/Interior B.C. (continued)
Skulls (text extract)
2.2. Skull measurements
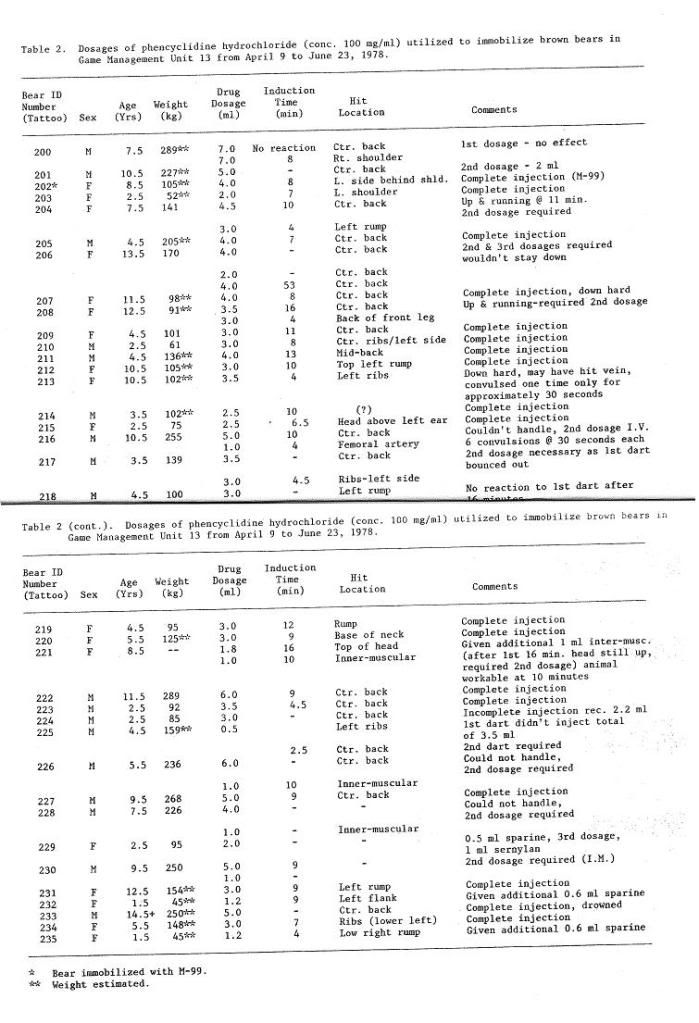
The removal of teeth from every live animal captured, and the problems inherent in decalcifying, sectioning and preparing so many sections, made the technique impractical for normal use in determining age of grizzly bears. Consequently, the use of morphological attributes for age determination was investigated.
Marks and Erickson (1966), using known-age black bears, showed that after the 8th year only the suture between the zygomatic process of the maxilla and the squamous portion of the temporals remained open, thereby allowing the skull to increase in width. Rausch (1963) and Mundy and Fuller (1964) demonstrated that the age of grizzly bears as determined by cementum rings is positively correlated with the zygomatic breadth of the cleaned skull.
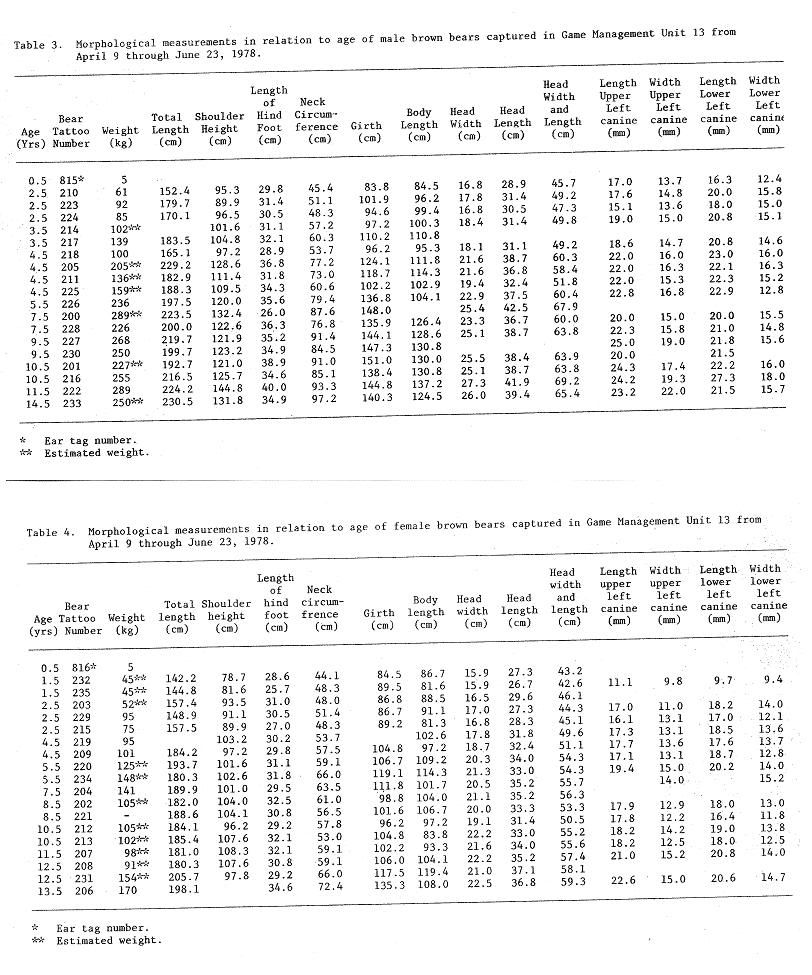
Mundy and Fuller (1964) combined measurements of male and female grizzlies for analysis. However, considerable sexual dimorphism was evident in body size of grizzly bears in the Yukon and it was thought that this difference might be reflected in skull size. Because the age determined from cementum annulations appeared to be accurate, a regression analysis was calculated from the comparison of this determination with the zygomatic breadth of the cleaned skull, or with the width of the skull in live animals. This was done for each sex separately. A correction factor was calculated to convert the live animal measurement to the equivalent breadth of the cleaned skull. During the study, 26 grizzly heads (19 males, 7 females) were measured for zygomatic breadth with the hide and flesh in place and again when the skulls were cleaned. The width of the fleshed and cleaned skulls averaged 94.4 per cent of the natural head width for both sexes. This figure was used as the correction factor in calculating regression lines.
Rausch (1963) elaborated on the variation in the condylobasal length of grizzly bear skulls in North America. In grizzlies found in the western United States and Canada, he demonstrated a decrease in condylobasal length from south to north. I hypothesized that within the Yukon Territory, or at least within the area of this study, there could be a decrease in the actual age-specific size of the skull from southern to northern Yukon and into the Northwest Territories. To determine whether inherent differences in relative skull dimensions existed within the population, the zygomatic breadth was plotted against the total skull length for all grizzlies measured during the study. No difference in relative skull size could be related to specific areas. There remained, however, differences in skull breadth relative to age of bears from different areas, and for that reason the total sample was divided into subsamples representing various regions for regression analyses of skull breadth and age. The skull material collected came from several areas that formed a semicircle north and east of the Central Yukon, and analyses of variance were run for the various areas.
For males, the samples from the Ogilvie, Wernecke, and Mackenzie Mountains showed no significant differences among themselves but each showed a significant difference from southern and central Yukon samples. However, the latter two samples were not significantly different from each other. The subsamples were grouped accordingly and two regression lines calculated for the male segment. The regression formulae for the two samples are:
1. Ogilvie, Wernecke, and Mackenzie Mountains
logy = 4.510008log* - 9.29264 n = 67, r = 0.922
2. southern and central Yukon logy = 3.95651 log* - 8.10861 n = 121, r = 0.924
The regression lines are shown in Figure 7. Male grizzlies from southern and central Yukon have a wider skull at a particular age than do animals from more northern locations in the Yukon or the Mackenzie Mountains. There is good evidence that the trend is reversed on the North Slope of the Yukon but the data have not yet been analyzed.
In the female segment no significant differences were found among any of the subsamples. All material was grouped and a single regression line calculated (Fig. 7).
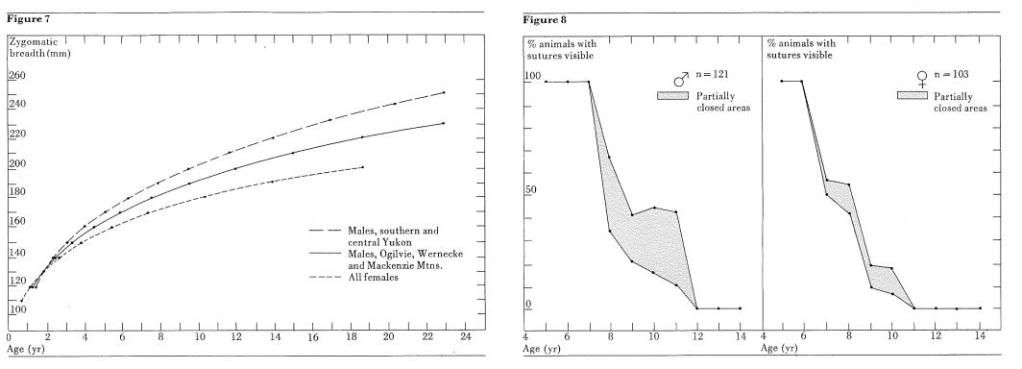
Table 8 shows the predicted ages for the above eight animals using firstly the equation derived from the entire sample, secondly the subgroup equations and finally the actual age as derived from the annuli in the cementum of the teeth. Although a more accurate prediction of the age of grizzly bears under 10 years of age can be obtained by using the subgroup equations, particularly around 10 years of age, this advantage is offset by a very large error found in older animals. It is concluded that the formula for the entire sample provides a more suitable basis on which to predict the age of a grizzly bear from the zygomatic breadth.
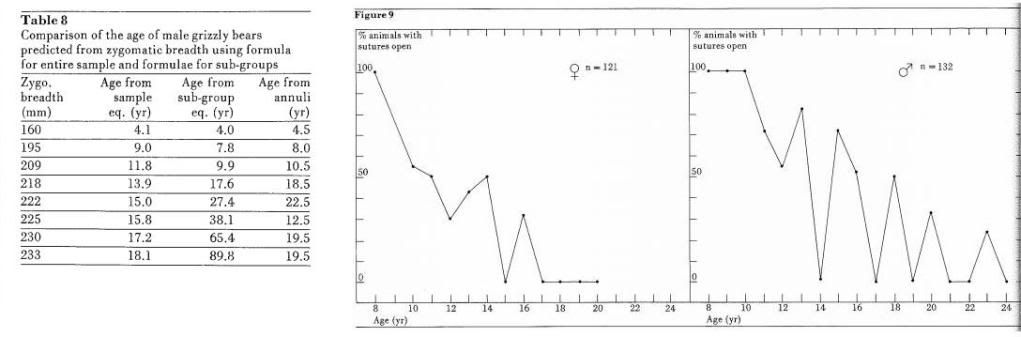
We carried out analysis on other skull measurements and on suture closure to assess their usefulness in determining the 9- to 11-year class and obliterated in the 12 +-year animals. Marks and Erickson (1966) found that, in female black bears in Michigan, the animal was at least 4.5 years of age if the basioccipital-basisphenoid suture was completely obliterated. Grizzly skulls were examined and classified as to whether the basioccipital-basisphenoid suture was open, closed or partially closed. Figure 8 shows the percentage of animals in each category for various age classes. The sexes are treated separately. It can be seen that for males no animals in the 8th year had closed or partially closed basioccipital-basisphenoid sutures. At the same time no mind that the characteristic lasted until he 4th year. No skulls from animals over I years showed the feature. The sagittal rest developed with age, being absent in animals under 5 years. Old animals showed greater ossification of the crest but no objective measure could be found that correlated accurately with age. Marks and Erickson (1966) showed that the internasal and nasofrontal sutures closed as black bears aged. They found the earliest obliteration in females at 7.8 years of age and in males at 6.8 years. The grizzly skulls were classed as having sutures obliterated or open and the feature was correlated with age from cementum annulations. Figure 9 hows the percentage of skulls having open sutures for various age classes. In males, io sutures were obliterated in animals in he 11th year or younger. One male of 23 ears showed open sutures. In the females, no sutures were obliterated in the 9th year or younger and the oldest animal with an open suture was 16 years of age. The usefulness of the progression of closure of the internasal and nasofrontal suture is limited by the great variability within the population. A male grizzly with the sutures closed can be classed as 12 years or older and the female as 10 years or older. Nothing can be said definitely at the other extreme.
Source: Pearson, 1975.
Skulls (text extract)
2.2. Skull measurements
The removal of teeth from every live animal captured, and the problems inherent in decalcifying, sectioning and preparing so many sections, made the technique impractical for normal use in determining age of grizzly bears. Consequently, the use of morphological attributes for age determination was investigated.
Marks and Erickson (1966), using known-age black bears, showed that after the 8th year only the suture between the zygomatic process of the maxilla and the squamous portion of the temporals remained open, thereby allowing the skull to increase in width. Rausch (1963) and Mundy and Fuller (1964) demonstrated that the age of grizzly bears as determined by cementum rings is positively correlated with the zygomatic breadth of the cleaned skull.
Mundy and Fuller (1964) combined measurements of male and female grizzlies for analysis. However, considerable sexual dimorphism was evident in body size of grizzly bears in the Yukon and it was thought that this difference might be reflected in skull size. Because the age determined from cementum annulations appeared to be accurate, a regression analysis was calculated from the comparison of this determination with the zygomatic breadth of the cleaned skull, or with the width of the skull in live animals. This was done for each sex separately. A correction factor was calculated to convert the live animal measurement to the equivalent breadth of the cleaned skull. During the study, 26 grizzly heads (19 males, 7 females) were measured for zygomatic breadth with the hide and flesh in place and again when the skulls were cleaned. The width of the fleshed and cleaned skulls averaged 94.4 per cent of the natural head width for both sexes. This figure was used as the correction factor in calculating regression lines.
Rausch (1963) elaborated on the variation in the condylobasal length of grizzly bear skulls in North America. In grizzlies found in the western United States and Canada, he demonstrated a decrease in condylobasal length from south to north. I hypothesized that within the Yukon Territory, or at least within the area of this study, there could be a decrease in the actual age-specific size of the skull from southern to northern Yukon and into the Northwest Territories. To determine whether inherent differences in relative skull dimensions existed within the population, the zygomatic breadth was plotted against the total skull length for all grizzlies measured during the study. No difference in relative skull size could be related to specific areas. There remained, however, differences in skull breadth relative to age of bears from different areas, and for that reason the total sample was divided into subsamples representing various regions for regression analyses of skull breadth and age. The skull material collected came from several areas that formed a semicircle north and east of the Central Yukon, and analyses of variance were run for the various areas.
For males, the samples from the Ogilvie, Wernecke, and Mackenzie Mountains showed no significant differences among themselves but each showed a significant difference from southern and central Yukon samples. However, the latter two samples were not significantly different from each other. The subsamples were grouped accordingly and two regression lines calculated for the male segment. The regression formulae for the two samples are:
1. Ogilvie, Wernecke, and Mackenzie Mountains
logy = 4.510008log* - 9.29264 n = 67, r = 0.922
2. southern and central Yukon logy = 3.95651 log* - 8.10861 n = 121, r = 0.924
The regression lines are shown in Figure 7. Male grizzlies from southern and central Yukon have a wider skull at a particular age than do animals from more northern locations in the Yukon or the Mackenzie Mountains. There is good evidence that the trend is reversed on the North Slope of the Yukon but the data have not yet been analyzed.
In the female segment no significant differences were found among any of the subsamples. All material was grouped and a single regression line calculated (Fig. 7).
Table 8 shows the predicted ages for the above eight animals using firstly the equation derived from the entire sample, secondly the subgroup equations and finally the actual age as derived from the annuli in the cementum of the teeth. Although a more accurate prediction of the age of grizzly bears under 10 years of age can be obtained by using the subgroup equations, particularly around 10 years of age, this advantage is offset by a very large error found in older animals. It is concluded that the formula for the entire sample provides a more suitable basis on which to predict the age of a grizzly bear from the zygomatic breadth.
We carried out analysis on other skull measurements and on suture closure to assess their usefulness in determining the 9- to 11-year class and obliterated in the 12 +-year animals. Marks and Erickson (1966) found that, in female black bears in Michigan, the animal was at least 4.5 years of age if the basioccipital-basisphenoid suture was completely obliterated. Grizzly skulls were examined and classified as to whether the basioccipital-basisphenoid suture was open, closed or partially closed. Figure 8 shows the percentage of animals in each category for various age classes. The sexes are treated separately. It can be seen that for males no animals in the 8th year had closed or partially closed basioccipital-basisphenoid sutures. At the same time no mind that the characteristic lasted until he 4th year. No skulls from animals over I years showed the feature. The sagittal rest developed with age, being absent in animals under 5 years. Old animals showed greater ossification of the crest but no objective measure could be found that correlated accurately with age. Marks and Erickson (1966) showed that the internasal and nasofrontal sutures closed as black bears aged. They found the earliest obliteration in females at 7.8 years of age and in males at 6.8 years. The grizzly skulls were classed as having sutures obliterated or open and the feature was correlated with age from cementum annulations. Figure 9 hows the percentage of skulls having open sutures for various age classes. In males, io sutures were obliterated in animals in he 11th year or younger. One male of 23 ears showed open sutures. In the females, no sutures were obliterated in the 9th year or younger and the oldest animal with an open suture was 16 years of age. The usefulness of the progression of closure of the internasal and nasofrontal suture is limited by the great variability within the population. A male grizzly with the sutures closed can be classed as 12 years or older and the female as 10 years or older. Nothing can be said definitely at the other extreme.
Source: Pearson, 1975.