|
|
Post by grrraaahhh on Apr 15, 2011 6:59:28 GMT -9
|
|
|
|
Post by grrraaahhh on Apr 15, 2011 7:00:24 GMT -9
Vegetation maps for North America and Greenland:a-c) Percent land surface covered by a) woody vegetation, b) needleleaved trees and shrubs, and c) broadleaved trees and shrubs (DeFries et al. 1999b, DeFries et al. 2000); d) the current distribution of vegetation formations, based on the International Geosphere-Biosphere Programme (IGBP) classification scheme (Loveland et al. 2000); and e) potential vegetation, representing the likely state of vegetation prior to European settlement (Fedorova et al. 1990). 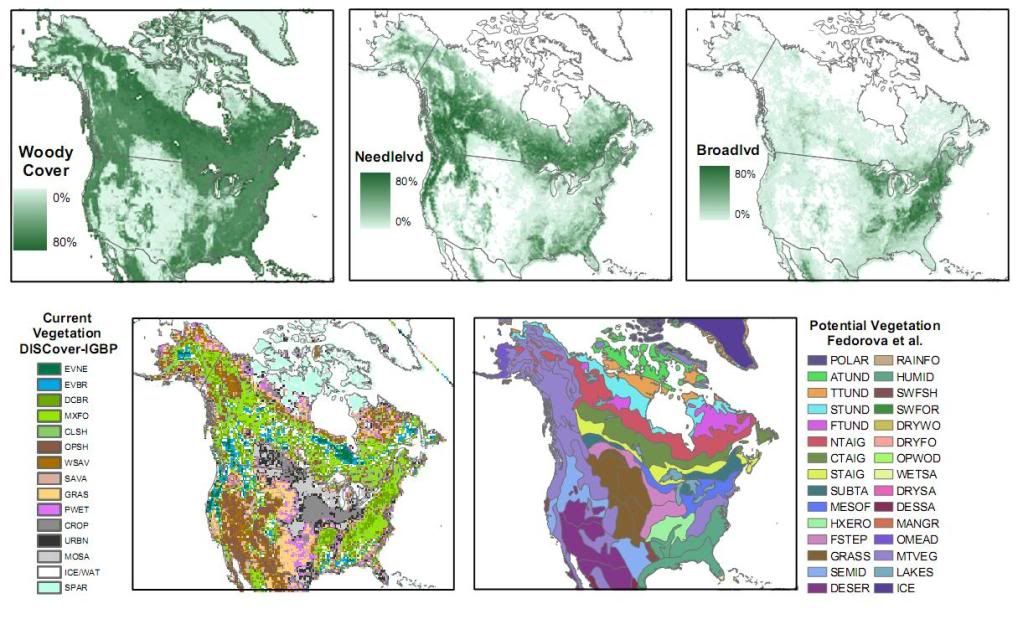 Vegetation codes for the DISCover vegetation map are as follows: Vegetation codes for the DISCover vegetation map are as follows:EVNE= Evergreen Needleleaf Forest, EVBR= Evergreen Broadleaf Forest, DCBR= Deciduous Broadleaf Forest, MXFO= Mixed Forest, CLSH= Closed Shrublands, OPEN= Open Shrublands, WSAV= Woody Savannas, SAVA= Savannas, GRAS= Grasslands, PWET= Permanent Wetlands, CROP= Croplands, URBN= Urban and Built-Up, MOSA= Cropland/Natural Vegetation Mosaic, ICE= Snow and Ice, and SPAR= Barren or Sparsely Vegetated. Vegetation codes for the Fedorova et al. (1990) map are:POLAR= Polar deserts, ATUND= Arctic tundra, TTUND= Typical tundra, STUND= Southern tundra, FTUND= Forest-tundra, NTAIG= Northern taiga, CTAIG= Central taiga, STAIG= Southern taiga, SUBTA= Subtaiga, MESOF= Mesophytic forests, HXERO= Hemixerophytic forests, FSTEP= Forest-steppes, GRASS= Grasslands, SEMID= Semideserts, DESER= Deserts, RAINFO= Rain forests, HUMID= Humid forests, SWFSH= Seasonally wet forests and shrublands, SWFOR= Seasonally wet forests, DRYWO= Dry forests, woodlands, and shrublands, OPWOD= Open woodlands, WETSA= Wet savannas, DRYSA= Dry savannas, DESSA= Desert-like savannas, MANGR= Mangrove, OMEAD= Oceanic meadows, MTVEG= Mountain vegetation, LAKES= Lakes/Reservoirs, ICE= Glaciers. 
|
|
|
|
Post by grrraaahhh on Apr 15, 2011 21:13:13 GMT -9
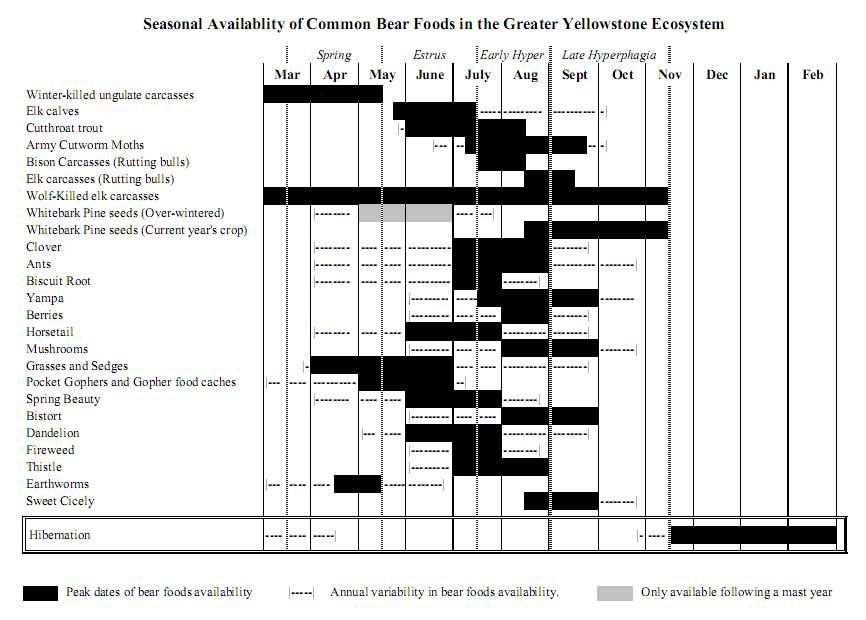
The Grizzly Bears of Yellowstone National Park (Quick Recap) History History A heated debate began in the 1960s and 1970s and grew to a national scope concerning the grizzly bears in the GYE. For decades, grizzly bears were allowed to rummage through garbage dumps searching for food. As early as the 1940s, some researchers suggested closing the open-pit dumps within Yellowstone National Park. In 1963, the Advisory Board on Wildlife Management in the National Parks released the “Leopold Report” which recommended that natural ecosystems should be recreated, including predator/prey relationships.  By 1967, Yellowstone National Park’s Superintendent Anderson began to implement recommendations of the Leopold Report. The Park began closing the open-pit dumps and bears were to be weaned off garbage. Some researchers suggested gradually phasing out the dumps, but the Park staff closed everything, rationalizing that there were enough backcountry bears that did not use dumps to sustain the mortality. The controversy continued because grizzly bear mortality increased substantially as dumps were closed. Between 1967 and 1972, a minimum of 229 Yellowstone ecosystem grizzlies died. The IGBST was formed by the Department of Interior in 1973 as a direct result of this controversy. The high mortality that followed dump closure and concerns for the population’s future led to its listing as threatened under the Endangered Species Act in 1975. Early research by the team indicated that following listing, the population continued to decline into the 1980s. This information was the foundation and impetus for the formation of the Interagency Grizzly Bear Committee (IGBC) in 1983. The IGBC, represented by administrators from federal and state agencies, implemented several regulations on federal lands designed to reduce human-caused grizzly bear mortality. These management policies, in concert with favorable environmental conditions, halted the population’s decline. Grizzly bear numbers have increased since the mid-1980s and today bears again occupy historical range well beyond Yellowstone National Park. USGS SOURCE LINK: www.nrmsc.usgs.gov/research/igbst-home.htm#HistoryTo be continued.....
|
|
|
|
Post by grrraaahhh on Apr 16, 2011 8:49:50 GMT -9
|
|
|
|
Post by grrraaahhh on Apr 16, 2011 10:47:24 GMT -9
Yellowstone National Park (continued)Weight & MeasurementsI. 1959-1970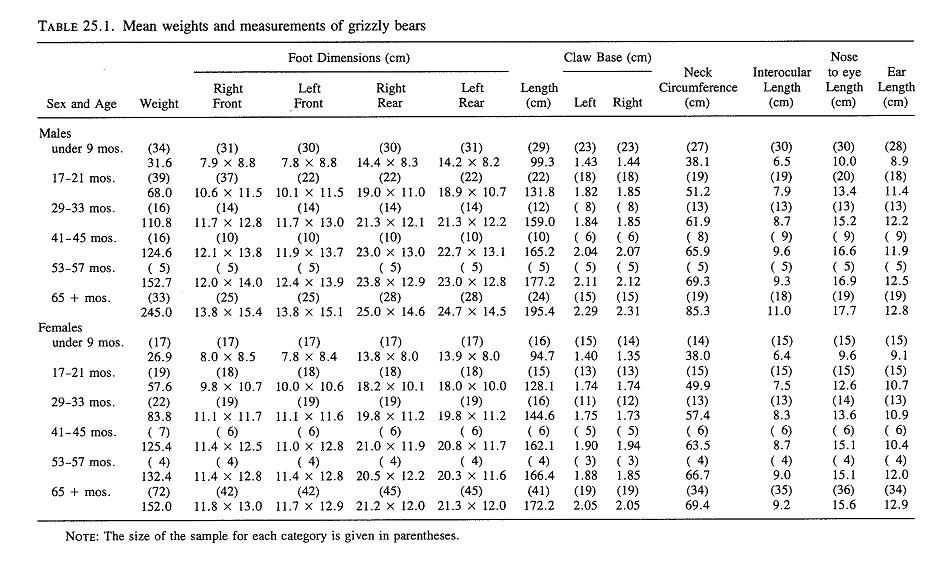 Text Extract: "Reliable data on measurements of grizzly bears exist for populations in the Brooks Range of Alaska (Rausch 1963), the Yukon Territory (Pearson 1975), and the Yellowstone ecosystem (Craighead and Craighead 1973b). Weights of large adult animals from the Yellowstone ecosystem ranged from 158 to 204 kg for females, and from 363 to 500 kg for males. Average weights and physical dimensions by age classes of Yellowstone grizzly bears are given in table 25.1 (J. J. Craighead unpublished data)." Craighead, J. J., and J. A. Mitchell. 1982. Grizzly Bear. Pp. 515-556 in J. A. Chapman and G. A. Feldhamer (eds.). Wild Mammals of North America: biology, management and economics. John Hopkins Univ. Press, Baltimore, MD. II."Blanchard (in press) found that in Yellowstone National Park, male grizzlies were consistently heavier than females for all age classes except cubs and yearlings. Sexual dimorphism beginning at age 2 was also apparent in other study areas (Troyer and Hensel 1969, Pearson 1975). Males in Yellowstone steadily gained weight until at least 15 years, but the mean annual rate of weight increase for males aged 4-15 (5.6%) was much less than the rate for cubs through 3 years (42.1%). The mean weight of adult males was 29% greater than for females and body measurements were 8-17% greater. Males attained full size in 7 of 11 body measurements by age 6 and in all 11 by age 9." SOURCE: Grizzly Bear Compendium (1987) by the Inter agency Grizzly Bear Committee. 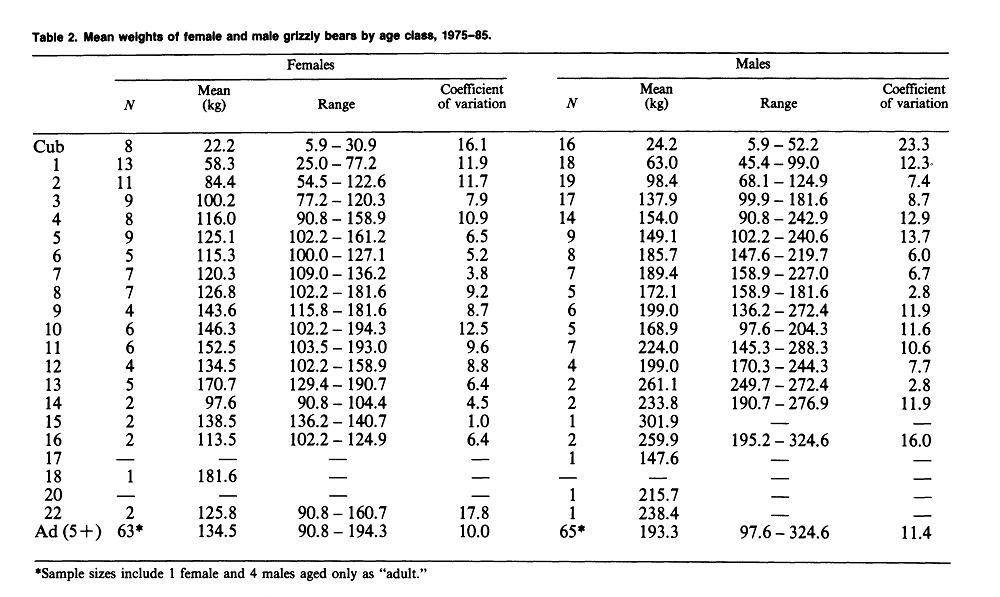 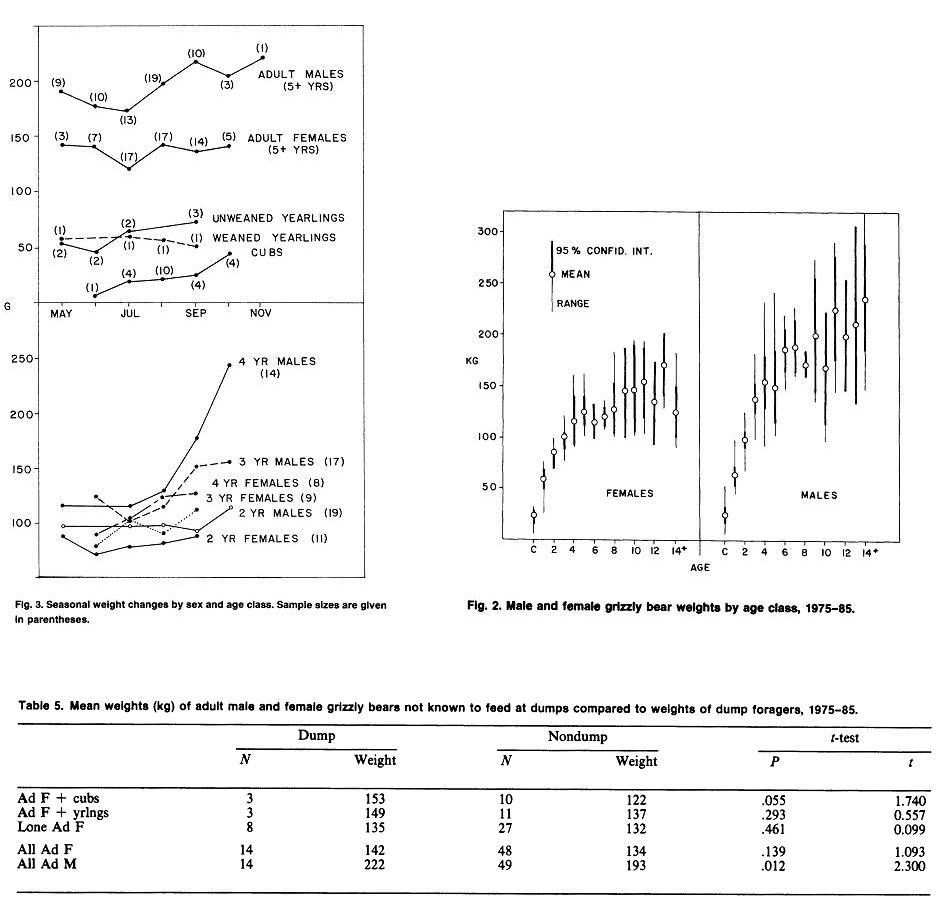 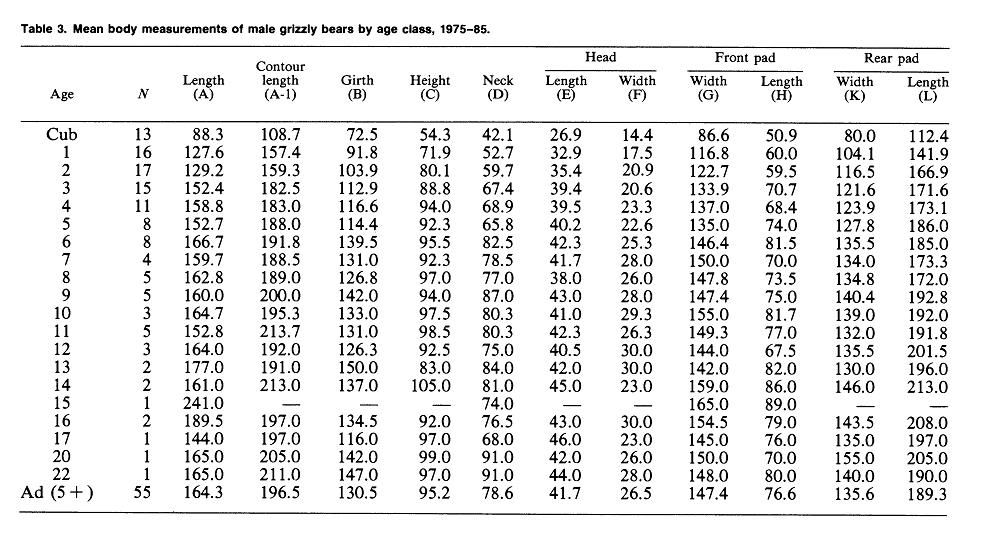 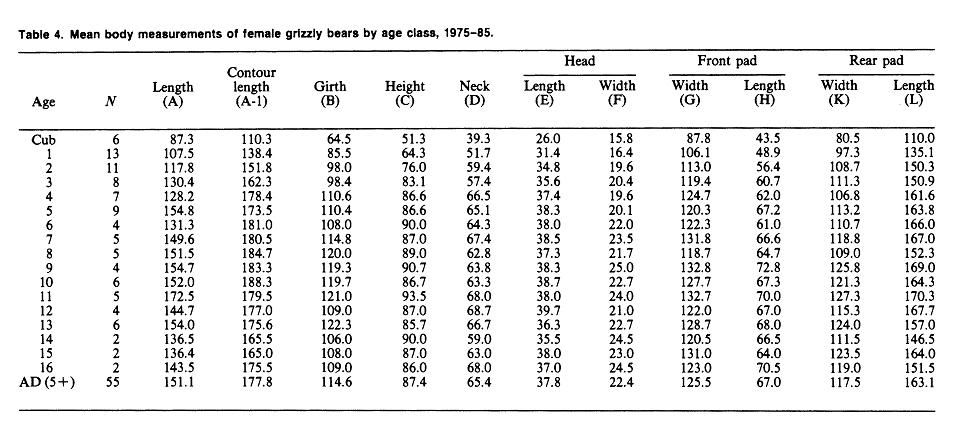 Blanchard, B. 1987. Size and growth patterns of the Yellowstone grizzly bear. International Conference on Bear Research and Management 7:99-107. PDF LINK: www.google.com/url?q=http://www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_7/Blanchard_Vol_7.pdf&sa=U&ei=-tapTeOSH47EsAOus8j5DA&ved=0CBYQFjAA&usg=AFQjCNEX0N4nRyQtzC8ZznyXgIlw9pIIlA
|
|
|
|
Post by grrraaahhh on Apr 16, 2011 10:49:07 GMT -9
Yellowstone National Park (continued)Book Extract: AGGRESSIVE BEHAVIORS OF ADULT MALES AT THE TROUT CREEK ECOCENTERAggressive encounters among aggregation members frequently involved spectacular feats of skill, strength, and endurance. Descriptions of these from abridged field notes for the year 1963 give some sense of the atmosphere of aggressive power that created the hierarchy each season. Marked male bears are identified by a number and/or a name; unmarked but individually recognizable bears are referred to by name only. Journal notes are as follows: 18 June 1963—Scarneck has not yet been sighted in the aggregation. Short-eared Boar is closely attending female No. 112. Inge (No. 12) is following female No. 65; she is in estrus. Inge is leaving No. 65 to challenge Short-eared Boar. He pursues him over a quarter mile [0.4 km]; Short-eared Boar turns and makes a stand. Inge closes with his opponent, parries, and jockeys for position. They move cautiously like wrestlers seeking advantage and leverage. Inge suddenly bores in, seizes Short-eared Boar by the throat, leveraging his body in a twisting turn. This maneuver throws the 750-pound male to the ground on his back. We can hear the two roaring from well over a third of a mile [0.5 km] away. Short-eared Boar regains his feet; the two stand, man-like, their jaws locked, striking fierce blows with powerful forepaws. They relinquish their holds and drop to all fours. Inge makes a bluffing charge but Short-eared Boar does not retreat. They again lock jaws and Inge forces Short-eared to back up. It is apparently not a concession, but a maneuver. Short-eared lunges for Inge's throat. Inge lowers his head and dives for the Short-eared Boar's groin. They roll and tussle. Neither can get a firm hold. Inge keeps the pressure on, driving steadily on after each lunge, backing Short-eared down the slope some 20 feet [6.1 m]. Short-eared turns and runs; he is pursued by Inge at a fast pace through the sage for half a mile [0.8 km]. Inge stands, watching his retreating contender for several minutes. He ambles slowly back to female No. 65 and together they move over the hill, the male nudging the female along until they disappear from sight.
Short-eared Boar circles back to the dump and immediately shows interest in female No. 40, sniffing her genitalia (checking). The other males have shown no interest in her this evening. Short-eared mounts No. 40 and breeds her. Number 40 is receptive and the copulation lasts 19 minutes. Number 40 moves to a feeding site; Short-eared Boar moves to the creek, enters the water, and lays down. Female No. 40 was bred earlier by No. 88 [Patch-eye] on 11 June and by Inge on 14 June. Today's breeding extends her observed estrous period to a minimum of eight days. Inge appears to be the most dominant of the large males, but he is not yet recognized as alpha. The Grizzled Boar appears to be Inge's chief contender, but he has not yet appeared today.
19 June 1963—Female No. 112 is receptive to male No. 88, but he has been unable to breed her. Fighting and chasing contenders has interrupted his advances. Female No. 65 is present and apparently still in estrus as she is attracting the large, aggressive males. Inge is alert, his eyes fastened on male No. 88, 40 yards [36.5 m] away. He appears oblivious to female No. 65 and to other bears around him who are also alert and moving outward to a fringe of nervous animals. With ears erect and head thrust forward, Inge stands motionless, watching. In prelude to battle, each male approaches the other in a slow stiff-legged walk. Inge stomps his hind feet alternately downward and briefly holds his legs rigid before taking the next step in a similar manner. Advancing in this swaggering, robotlike gait, the males move closer. Both animals salivate profusely and urinate. The approach and ultimate showdown is delayed by frequent pauses as the contenders eye one another. Each appears to look for an appeasement sign or a propitious moment to charge. Inge continues his stiff-legged walk toward No. 88 and pauses. Number 88 moves in similar manner toward Inge. Both bears are studies in concentration: both seem oblivious to their surroundings. The pace increases. They are now 50 feet [15 m] apart. Most members of the aggregation have stopped feeding and are facing the contenders. Emitting a low, guttural growl, ears laid back and head low, Inge charges. It is amazingly fast. We detect no warning signal but No. 88 apparently does, for in split seconds he is prepared to defend himself. He does not meet Inge face-on as he has other contending males. He wheels around, and as he does so, Inge's jaws close on his rump, tearing free a 3-inch [7-cm] flap of hide exposing a white slash. Without losing stride or momentum, Inge climbs over No. 88, breaking him down. Number 88 twists under Inge's weight and rolls to his back, with all four feet fending off his opponent. Inge sinks his teeth deep into No. 88's groin as he lies pinned on his back. There is a brief tussle, the roaring loud and continuous. Inge rears back, lifting his head. As he does so, he lifts the near-700-pound [320-kg] bear clear of the ground and shakes him. The action is occurring in seconds, and I find it hard to believe my eyes. Both bears hit the ground in a tumbling roll. Number 88 has Inge by the neck as he finally regains his feet. He releases his hold and clamps on Inge's jowls, shaking his massive head. Inge rears back, fighting free at the expense of a ripped and bleeding lower jaw. The two stand erect face to face, jaws gaping, teeth bared, emitting rumbling growls. They drop to all fours, and in a synchronous lunge rise together to clash teeth, slap with fore-paws, and engage and disengage their jaws as each seeks an advantage. Inge throws his weight to the right, attempting to twist No. 88 to the ground, but No. 88 counters with a lunge to the left, his teeth imbedded in Inge's neck. They break apart and silently eye one another. The pause in action is only a few seconds but seems longer. Inge is the first to move. He steps upward to the right, taking advantage of the slope. Number 88 makes a leg dive; Inge sidesteps and slaps No. 88 across the shoulder with a terrific swipe of his right paw. I hear the slap and see deep furrows ripple through the fur. Number 88 lurches backward from the blow and Inge drives forward for No. 88's throat, exposed for a fraction of a second as he seeks his balance. Rising on hind legs, the two embrace with an audible clash of teeth. Biting and swiping, they exchange roars that carry across the valley and noticeably excite the spectator bears. The roaring subsides to growling and the contestants, now only feet apart, stand motionless eyeing one another with steady stares. Neither moves in this control of wills; 88 does not retreat but slowly lowers his head to one side, and eye contact is momentarily broken. Inge's head goes up, his ears erect. Number 88 holds his submissive pose, but with eyes on Inge; still, he does not retreat. Inge deliberately turns his back on 88 and very slowly moves 60 feet [18 m] to claim female No. 65. Number 88 could attack Inge from behind but he does not. This recognition of defeat is never breached. The contest is over; for this day Inge is victorious. There will be no sneak attack, no renewal of hostilities. The other bears begin to feed as though suddenly released. I can almost feel the tension subside. Number 65 had been receiving the attention of the Large Brown Boar while Inge was fighting. This male attempts to mount the female but retreats as Inge approaches. Number 88 remains where he had fought, breathing heavily, then climbs the embankment and mixes with the other males now actively feeding. He moves on to the creek. Inge, his chest expanding and contracting like huge bellows, stands with female No. 65 at his side, eyeing the aggregation. It is a challenging pose, but no bear responds. Ten minutes elapse before Inge moves. He appears to have lost interest in the female but his presence keeps other males at a distance. Neither Inge nor No. 88 appears to be seriously injured although both show wounds from the conflict. Inge is clearly dominant over No. 88 this day, but he is not yet recognized as alpha. There are other males yet to contend with.
26 June 1963—Inge (No. 12) and the Grizzled Boar [never captured and marked] have had three encounters earlier in the season, but none have been decisive. Since then, Inge and the Grizzled Boar have avoided one another. When they have approached closely, Inge has appeared to dominate. The Grizzled Boar has not challenged Inge and Inge has been relatively tolerant of his presence. The Grizzled Boar has had successful encounters with Short-eared Boar and Scarneck. He is powerful and aggressive but keeps his distance from Inge. There are three females present that are now in estrus. Inge has been aggressive toward all bears this evening, but there have been no battles.
Inge (No. 12) is now moving among the 60 bears present. All show him deference—some by moving aside, others by lowering the head, and some by ignoring him completely. The Grizzled Boar and the Short-eared Boar are squared off, intently eyeing one another. Inge moves swiftly toward them, attacking the Grizzled Boar from behind. The Grizzled Boar is unaware of his approach until he is crushed to the ground. Inge does not carry the fight but allows the Grizzled Boar to retreat. [Inge] has not been feeding; he seldom does. He is patrolling the aggregation, exerting authority. The Grizzled Boar moves toward a choice feeding site, scattering younger bears and female No. 96 with her one cub. A growl and a rapid step is all it takes for,him to clear the site. The Grizzled Boar has fresh scars on his jowls and neck that were not there a day ago; he has been in a battle we did not observe, perhaps in the backcountry. His behavior toward Inge has changed drastically. He is now relatively submissive. He probably fought Inge for female No. 65 or possibly for female No. 40. In any case, he and Inge have battled, and Inge is now the alpha male. The Grizzled Boar recognizes this, as does the entire aggregation. There was no need to have seen the showdown; evidence speaks in the actions and behaviors of every bear. All defer to the "boss," from weaned yearlings and 2-year-olds to Inge's other major contenders, Short-eared Boar, No. 88, and Scarneck. Inge has dominated all the large aggressive males and has reclaimed his former status, relinquished to Scarneck last season (1962). We might say that he has established himself as alpha in 26 days of aggressive action and constant vigilance, or that the aggregation has accepted him as the dominant male after a 26-day period of contention. He has vigilantly checked the fertility of estrous females and has copulated at least three times with two different ones. He has earned the right to select females without a battle and to take choice feeding sites throughout the summer.Encounters such as those just described occurred each year and determined the ascension and succession of alpha males from one year to the next. The organization of the remainder of the male hierarchy fell into place by the same process. Source: The Grizzly Bears of Yellowstone: their ecology in the Yellowstone ecosystem, Craighead, J.J. (1995). shaggygod.proboards.com/index.cgi?board=horribilis&action=display&thread=411
|
|
|
|
Post by grrraaahhh on Apr 16, 2011 19:12:50 GMT -9
|
|
|
|
Post by grrraaahhh on Apr 17, 2011 15:32:32 GMT -9
Glacier National Park (Montana) Abstract Bear management in Glacier National Park (GNP), Montana, has evolved from 1960 to 1994. Grizzly bears (Ursus arctos) and black bears (U. americanus) have become more highly valued in both a social and an ecological context. Management has shifted from focusing on removing problem animals to preventing problem bear behavior by modifying human behavior. Reducing the availability of human foods to wildlife and tolerating natural defensive reactions by bears coincided with a decline in the number of bears killed or relocated. Reported sightings of black and grizzly bears have risen from 192 in 1967 to 2,075 in 1994, suggesting that bear populations have increased. However, we suggest the number of bear sightings is an unreliable indicator of population trend. Human injuries resulting from black bear encounters declined to near zero with the control of human food and garbage. The grizzly bear-inflicted human injury rate, however, continued to increase. Progress in understanding the causes will not be made until reliable information on bear populations and human backcountry use is available. A summary of bear management in Glacier National Park, Montana, 1960-1994.PDF LINK: www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_10/Gniadek_Kendall_Vol_10.pdf Annual Food Habits of Bears:Proportionate contribution of major food classes to the total annual diet volume of bears in Glacier National Park as determined by fecal analysis, 1967-71 and 1982-85 (n=1514)  Seasonal food habits: Proportionate contribution of major food classes to seasonal diet volume of bears in Glacier National Park 1982-88. LIMITED TEXT EXTRACT:Power of Sign Surveys to Detect Bear Population Trends: The grizzly bears in Glacier National Park represent the keystone of the Northern Continental Divide Ecosystem population in NW Montana. Because of its proximity to Canadian bear populations, large land area, and high proportion of designated wilderness and national park lands, the Northern Continental Divide Ecosystem has the best long-term prospects of supporting a viable grizzly bear population among the six areas designated as grizzly bear recovery zones in the United States. Yet there never has been a demographic study of the Glacier National Park grizzly bear population and no data exist on population size, status, or trend. The grizzly bear population in the Northern Continental Divide Ecosystem can not be delisted without reliable information on ecosystem-wide population trends, of which the bears in Glacier National Park are a critical component. Until now, statistically rigorous grizzly population studies in forested habitat could only be accomplished with radio telemetry. Using DNA to Monitor Grizzly Bear Population Trends: Recent advances in genetic technology allow identification of species, sex, and individuals from DNA extracted from bear hair and scats without handling bears. With proper survey design, identification of individuals and sex typing data can be used to determine (1) minimum population size, (2)population trends for both species, and (3) genetic diversity of the populations. Now that individual bears can be identified from hair and scats, sign surveys to monitor population trend will be more powerful than demonstrated by Kendall et al. (1992). In a study that began in 1998, DNA is being analyzed from bear sign collected along survey routes in Glacier National Park and adjoining lands. The fundamental goal of this work is to develop a nonintrusive, relatively inexpensive method to monitor grizzly bear population status and trend in the Northern Continental Divide Ecosystem. The number of individuals and species identified will be used to develop minimum counts and baseline population indices. The power of surveys with varying sample designs will be compared using methods described by Kendall et al. (1992). Grizzly bear conservation biology: An assessment of the status of grizzly bear populations in the contiguous United States offers a new approach to population viability analysis which has implications for grizzly bear conservation strategy (Mattson et al. 1995). A study, begun in 1998, that will use DNA extracted from sign to monitor bear population trends, will also address bear conservation genetics as described above. USGS LINK: www.nrmsc.usgs.gov/research/grizzlyb.htmFrom Local food habits of the grizzly bear in Montana (1986): Abstract Grizzly bear (Ursus arctos horribilis) scats were collected from 4 western Montana study areas from 1976 to 1979 to determine differences in food item selection. Fruit was important to grizzly bears in all areas although the species consumed and the apparent degree of use varied. Globe huckleberry (Vaccinium globulare) was important to grizzly bears in the North and South forks of the Flathead River but was rarely eaten in other areas. Domestic apples (Malus spp.) and plums (Prunus spp.) were eaten extensively by Mission Mountain grizzly bears. Grasses and sedges were a staple food to bears in all areas; variable use of Umbelliferae was found. The nuts of whitebark pine (Pinus albicaulis) were eaten extensively by East Front grizzly bears only, and biscuit-root (Lomatium spp.) roots were dug to varying degrees in all areas. Yellow hedysarum (Hedysarum sulphurescens) roots were an important spring and autumn food to North Fork grizzly bears only. Horsetails (Equisetum spp.), clover (Trifolium spp.), and dandelions (Taraxacum spp.) were important in all areas throughout the grizzly bears' active period. These data suggest that substantial local variation occurs in grizzly bear food habits in Montana. These differences should be considered in land management plans that call for maintenance or enhancement of grizzly bear habitat. PDF LINK: www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_6/Mace_Jonkel_Vol_6.pdf
|
|
|
|
Post by grrraaahhh on Apr 18, 2011 16:04:36 GMT -9
Glacier National Park (continued) Sow grizzly bear, GNP. Abstract We present the first rigorous estimate of grizzly bear (Ursus arctos) population density and distribution in and around Glacier National Park (GNP), Montana, USA. We used genetic analysis to identify individual bears from hair samples collected via 2 concurrent sampling methods: 1) systematically distributed, baited, barbed-wire hair traps and 2) unbaited bear rub trees found along trails. We used Huggins closed mixture models in Program MARK to estimate total population size and developed a method to account for heterogeneity caused by unequal access to rub trees. We corrected our estimate for lack of geographic closure using a new method that utilizes information from radiocollared bears and the distribution of bears captured with DNA sampling. Adjusted for closure, the average number of grizzly bears in our study area was 240.7 (95% CI = 202–303) in 1998 and 240.6 (95% CI = 205–304) in 2000. Average grizzly bear density was 30 bears/1,000 km2, with 2.4 times more bears detected per hair trap inside than outside GNP. We provide baseline information important for managing one of the few remaining populations of grizzlies in the contiguous United States. Grizzly Bear Density in Glacier National Park, Montana (PAID): www.csa.com/partners/viewrecord.php?requester=gs&collection=ENV&recid=8905536Abstract Vegetation was sampled on 330 sites known to be used by grizzly bears (Ursus arctos). The response to disturbance of 6 shrub species important as grizzly bear foods was determined by comparing their percent canopy cover on disturbed sites with that on undisturbed, old-growth sites. Overall, the canopy cover of these species was higher on sites burned by wildfire 35-70 years ago than on comparable old-growth sites. The canopy cover of these species was generally less on clearcut sites where the slash was bulldozer-piled than on burned sites. The shrub response on clearcut sites where slash was not treated was intermediate; some shrubs increased while others declined. Site treatment is at least partially responsible for this differential response; bulldozer-scarification apparently destroys the vegetative reproductive organs of these shrubs. Habitat use patterns of 4 radio-collared grizzly bears were studied in 1979. Grizzly bears preferred snowchutes, ridgetops, and creek bottoms during the spring; they preferred shrubfields, slabrock, ridgetops, and creek bottoms during the summer/fall. Cutting units and habitat affected by open, travelled roads were avoided throughout the active season. Cutting units used by grizzly bears were generally isolated from human disturbance factors and provided nearby cover (within 50 m) in the form of well-developed shrub strata, leave trees, and cutting unit boundaries. Logging and Wildfire Influence on Grizzly Bear Habitat in Northwestern Montana PDF LINK: www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_5/Zager_Jonkel_et_al_Vol_5.pdf
|
|
|
|
Post by grrraaahhh on Apr 22, 2011 8:02:54 GMT -9
|
|
|
|
Post by grrraaahhh on Apr 22, 2011 8:30:54 GMT -9
Southwest Yukon/Interior B.C. (continued) KNP. From A Case History of Grizzly Bear Management in the Slims River Area, Kluane National Park Reserve, Yukon: Abstract A management planning program for Kluane National Park Reserve was completed in 1980. A major decision was made to develop a public transit system in the Slims River Area to facilitate visitor access to a large valley glacier. The transit system was not built and the valley was managed as a backcountry hiking area for an interim period. Characteristics of grizzly bear-people conflicts were monitored from 1981 to 1987. Park staff and 2,603 registered overnight backcountry users recorded 503 grizzly observations. Observations of solitary bears increased from 40% of total bear observations in 1981 to 84% in 1987. Frequency of avoidance behavior by grizzlies decreased whereas apparent neutral and approach behaviors increased. Incidents defined as serious were infrequent from 1981 to 1984 (n = 3). Serious incidents sharply increased in 1985 (n = 10) and continued to be relatively frequent in 1986 (n = 6) and 1987 (n = 9). Serious incidents were categorized as close approach or charge (n = 10), pack robbing (n = 8), food cache robbing (n = 2) and disturbance of tent camps (n = 4), facilities (n = 3) and vehicles (n = 1). Management actions resulted in the death of 5 grizzlies, relocation of 5 grizzlies and area closures. Our analysis of relevant documents from 3 national park planning and management processes indicated that grizzly bears were not adequately treated in plans and environmental assessments for the Slims River Area because of emphasis on the proposed public transit system. The relationship between habituation of grizzlies to people and food conditioning was not recognized in management of the Slims River Area as a wilderness hiking area. We considered national park management processes to be valid tools for grizzly management provided they are implemented by trained, knowledgeable staff that apply adequate information before making decisions. PDF LINK: www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_8/Leonard_Breneman_Frey_8.pdf KNP. From GRIZZLY AND BLACK BEAR DENSITIES IN INTERIOR BRITISH COLUMBIA: ABSTRACT We used genetic identification of individuals and mark-recapture modeling to estimate bear (Ursus) population size in both a mountainous and interior plateau environment in central eastern British Columbia. We sampled 591 sites within a 9,452-km2 study area during June and July 2000 and detected grizzly bears (Ursus arctos) at 199 sites and black bears (U. americanus) at 316 sites. We identified 326 different grizzly bears using microsatellite profiling; there were more females than males in the sample (35M:65F). After correcting for the effects of closure bias, we estimated density to be 12 grizzly bears/1,000 km2 (95% confidence interval [CI] 7–28) on the plateau and 49/1,000 km2 (CI 43–59) in the mountains. These densities are considerably higher than those predicted by the habitat-based extrapolation method used for grizzly bear management in British Columbia. The incorporation of hunter harvest impacts to population size caused the greatest decline, and therefore discrepancy, in numbers between the habitat-based extrapolation and the DNA-based estimate. We also sampled grizzly bears along 47 km of stream during August 2000 when Chinook salmon (Oncorhynchus tshawytcha) were in the streams. We detected 15 different grizzly bears at the 10 sites we sampled, and estimated there were 21 grizzly bears (CI 16–44) present along these streams during the 2 weeks we sampled. Eleven of the 15 bears detected along the salmon streams had been previously detected during spring sampling and the movement distance detected for most bears was within the range expected given home range sizes in this area. However, 2 male bears made what appeared to be extra-territorial movements to the salmon streams (35 and 48 km). These streams historically have modest salmon runs, which highlight the importance of salmon to interior grizzly bears. Grizzly bear densities are higher on the west slopes of the Rocky Mountains, including the front ranges such as the Selkirk and Purcell Mountains, than on the east slopes of the Rockies and in boreal and sub-plains environments. Subpopulations in the Rocky Mountain west slopes that are discontinuous with surrounding populations have lower landscape scale density than populations further north. Black bear densities were 270 bears/1,000 km2 (CI 173–458) on the plateau and 100/1,000 km2 (CI 55-210) in the mountains. Spatial patterns in black bear densities appear to be opposite that of grizzly bears, with the highest densities occurring in boreal and sub-boreal plains environments and lower densities in mountainous environments. This observation may be more due to competition with grizzly bears than differences in habitat productivity. DNA survey methods are expensive; thus, there will likely always be a need to use habitat-based extrapolations to predict bear abundance across broad areas. The Fuhr-Demarchi method as used now needs to be modified so that it is objective, repeatable and testable. We suggest that managers should use DNA-derived population estimates in various regions as benchmarks to calibrate the Fuhr-Demarchi method. Within the Parsnip/Herrick study area, our analysis suggests that current grizzly bear harvest levels are only sustainable in the Hart Ranges. Because salmon appear to be an important food source for grizzly bears in the upper Fraser River basin region, consideration should be given to retaining access to all salmon streams for bears during resource planning. PDF LINK: www.env.gov.bc.ca/omineca/documents/parsnip_grizzly_inventory_final.pdfNEXT: Diet & Weight.
|
|
|
|
Post by grrraaahhh on Apr 25, 2011 8:16:50 GMT -9
Southwest Yukon/Interior B.C. (continued)Diet (Text Extract)PerspectiveFrom 1963 to 1972 a study of the ecology of the northern interior grizzly eco-type was carried out on a selected study area in southwestern Yukon Territory, Canada. The study was initiated by the CWS as a result of acknowledged concern over the continuing reduction of grizzly bear numbers and distribution, and the need for sound ecological data on which management decisions could be based. One of the terms of reference for the CWS during that era was to carry out long-term research on northern mammals. It was anticipated that results from this study would be most useful in comparison with information obtained from studies that were in progress in Montana and Alaska on two other eco-types of grizzly. The population studied was assumed to be representative of the ecotype inhabiting the extensive, relatively dry and nonproductive interior mountains of northern B.C., Yukon, NWT, and Alaska. A study area was chosen that did not reflect a serious coastal influence (presence of spawning fish runs). It also was in an area that afforded complete protection from hunting and the population was thus assumed to be self-regulatory. Animals were captured and individually marked. Ecological data were collected from subsequent observations of animals that could be identified by sex and age. Radio-telemetry was also used to locate animals. It is hoped that the results will be useful to wildlife agencies, particularly the Yukon Territory Department of Game, in formulation of wise management programs for the grizzly bear.  Food habits 1.1. Introduction Food habits 1.1. IntroductionOne of the first aspects of grizzly bear populations usually investigated by research workers is the food habits of the ecotype in question. Information has been published on Montana (Martinka 1972), interior mountains of British Columbia Mundy 1963), Kodiak Island (Clark 1957), interior mountains of Alaska (Murie 1944), Kamchatka Province of USSR (Ostroumov 1966, 1968a, 1968b; Lobachev 1966), Mountains of northeastern Siberia (Kist-hinskii 1972), Baykal region of eastern Siberia (Ustinov 1960, 1965), Leningrad region of USSR (Sokov 1969), the Trenlino trovince of the Italian Alps (Krott 1962), Albruzzo National Park, Italy (Zunino and Herrero 1972), and Sweden (Haglund 1968). From a review of these studies, some general conclusions can be drawn. In no ilea where the grizzly bear has been studied does the species feed exclusively on one articular item throughout the annual cycle. Food items important in the early spring are often insignificant later in the season. The grizzly bear can utilize a wide election of foods ranging from a nearly completely herbivorous diet to a heavy dependence on animal matter. Thus the diet varies greatly from one geographical area to another and within areas, depending upon the season and the relative abundance of various food items. Most workers reported that grizzly bears fed heavily on vegetable matter immediately after emergence from the den in he early spring, although the actual type of plant varied from one area to another. Novikoveia/. (1969) reported that bears fed heavily on aspen leaves (Popidus spp.), Sokov (1969) found bears eating fennel shoots, (Foeniculum vulgurc) while others reported predominant use of grasses and roots (Clark 1957; Murie 1944; Mundy 1963; Martinka 1972; Ostroumov 1966; Ustinov 1960, 1965). However, interspersed in the literature are references to other food items used locally by grizzly bears during the spring to supplement or supplant the vegetable diet for short periods. Ustinov (1960, 1965) observed brown bears along the shore of Lake Baykal that fed almost exclusively on newly emerged caddis flies and fresh roe of gobies or chub for 7-10 days in June. Krott (1962) reported grizzly bears feeding heavily in the spring on common frogs (Rana temporaria) where they were abundant in some swamps in the Italian Alps. And Martinka (1972), in Glacier National Park, Montana, and Cole (1972) in Yellowstone National Park, Wyoming, found ungulate carrion, particularly elk (Cervus canadensis), an important food item for bears in the spring. The findings of our study present yet another variation in the feeding pattern adapted to fit the peculiarities of southwestern Yukon Territory. The results are based on 970 observations of grizzly bears feeding, from 1964 through 1969. Additionally, 128 faeces were collected and the contents identified. This analysis is presented in Table 9.
|
|
|
|
Post by grrraaahhh on Apr 25, 2011 8:19:16 GMT -9
Southwest Yukon/Interior B.C. (continued)Diet (continued)3.2. Results 3.2.1. Seasonal changes a. Spring (April-May)As bears emerged in early spring they first searched out open hill sides near the den. The favorite food was the roots of Hedysarum alpinum. Some animals remained on the alpine and subalpine meadows for the entire month of May, feeding mainly on Hedysarum but also on willow catkins (Salix spp.) and other green vegetation, particularly grasses, as it appeared. As the snow cover cleared, some animals moved down to the vegetated alluvial flats where Hedysarum grew in abundance. The choice of food items on these flats was the same as for alpine meadows. Although grizzly bears feed on carrion at this lime of year if it is available, only two cases were actually observed, both involving moose (Alcesalces gigas) that had been killed in spring snow-slides. b. Summer (June-July)During June, the bears disappeared from the open areas. During the early years of the study, it was thought that they were using the heavy spruce (Piceaglauca) forest, feeding on shrubs and herbs. During the last, two years, the animals were traced with r'adio-telcmelry to the upper edges of the forest and more importantly to the subalpine willow flats. Of the 108 observations of grizzlies in June 1969, 80 were recorded from the heavy willow zone above the timber line. Also, throughout the study, willow occurred most frequently in faeces collected in June or, in 1965, in late May (Table 9). As patches of grass appeared in the swales above limber, bears were observed grazing. Grass also occurred commonly in June faeces. Early in June bears were observed feeding on dry bear-berries and, although they did not appear in any of the faeces collected, this food item could be important locally. The pattern in early July was similar to June although the faeces analysis indicated greater feeding on grasses and slightly reduced feeding on willow catkins. This change may have been related to the emergence of lush green grasses at all altitudes. As the month progressed, bears began appearing along the lower edge of timber, moving in the transition zone between forest and alluvial flats. As the soapberries ripened (July 12-25), grizzlies were commonly observed browsing on the large patches growing in the transition zone and in the scrub zone on the alluvial flats. The immediate utilization of ripe soapberries is shown in the faeces analysis (Table 9). The early July sample showed four faeces (50 per cent of samples) containing soap berries; all four were collected between July 12 and 15. Nearly all faeces (17 of 20) collected during the latter half of July contained soapberries. The heavy use of soapberry habitat by grizzlies was evident from our observations of bears from the ground. Our activities (trapping and daily observations) were severely limited by accessibility, and as a result, our travel was mainly along the valley bottoms. There were 47 trap actions and 9 observations from the ground from July 1 to 15 over the 5-year period between 1965 and 1969 inclusive. The corresponding figures were 102 and 77 for July 16 to 31 for the same years. All but 15 of the 77 observations were of bears feeding on soapberries. We made an effort to locate grizzlies in other habitat types, perhaps feeding on other sources, but our complete lack of success indicates the relative insignificance of other food items at that time of year. Roots of Hedysarum, grasses, remains of wasps, and carrion were found in faeces collected during late July. The three faeces collected that did not contain soapberries were composed entirely of Hedysarum roots. These roots are found in abundance on the alluvial flats in the same areas as the soapberries. The preference for berries, if and when available, was supported by the fact that later in the season after the berries had ripened and disappeared, the bears frequented the same locations but fed almost exclusively on roots. 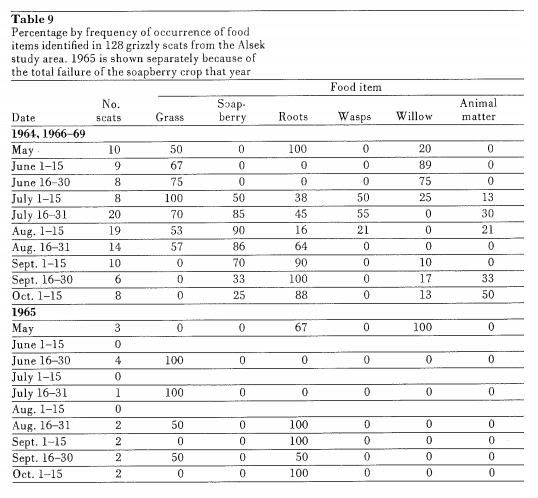 In 1965, there was a total failure of the soapberry crop. Grizzlies appeared at the' normal time in late July along the lower edges of the forest zone but no bear was seen more than once and the number of observations was drastically lower than in . the other years. Between July 15 and August 15 in 1965, we observed only 12 grizzlies (12 out of 139, or 8.6 per cent of the grizzly observations for the year). As a comparison, in 1966, 25 out of 102 (24.5 per cent) -and in 1967, 54 out of 132 (40.9 per cent) of the grizzly observations occurred during this period.
|
|
|
|
Post by grrraaahhh on Apr 25, 2011 8:20:16 GMT -9
Southwest Yukon/Interior B.C. (continued)
Diet (continued)
c. Fall (August-October)
During the first half of August, the bears continued feeding on soapberries along the alluvial flats. By mid-August, however, most of the berries had dried up or fallen from the bushes and the bears either moved up the creek bottoms into higher country, where the soapberry crop was later in maturing, or moved on to the more open alluvial islands and dug roots. With a few local variations, bears continue these activities until late September. Occasional concentrations of crowberry, blueberry (Vaccinium spp.) and bearberry in alpine regions attracted bears for short periods, although there were not sufficient quantities on the study area to have a major influence on the feeding pattern. However, wherever they did grow intensively, they were used heavily by grizzlies in September. For example, on September 5, 1969, 16 of 21 grizzlies observed during four hours of flying by helicopter, mainly in the area just south but including the southern and; eastern segment of the Alsek study area, were in the upper subalpine and alpine zone feeding on berries. Crowberry, and more frequently blueberry, occurred in denser patches down the Alsek River from the study area. The other five bears were feeding on roots within the study area.
In 1965, the year of complete soapberry crop failure, the bears appeared on the alluvial islands digging roots on August 20 and continued this activity; until at least October 4. In the July 15 - August 15 period of this year we recorded 12 out of 139 (8.6 per cent) observations, but the August 16 - September 15 period produced 65 out of 139 (46.8 per cent) of the year's observations.
With one variation, roots and late developing berries constituted the major food source until denning. During the year, bears were occasionally observed trying to dig out arctic ground squirrels (Spermo-philus undululatus) although in the rugged mountain terrain this was seldom successful. The ground squirrels, even when forced from their burrows, usually found cover in sticks before the bear could turn and capture them. In late September and October, however, when the alpine areas were covered with an even, fresh snow layer, bears ere commonly found traversing large areas digging for ground squirrels. Once the prey was forced from the burrow on to the surface of the snow the bears had good success. One bear was tracked on snow for four miles during which he dug out, captured, and ate at least seven ground squirrels. A further six attempts appeared to be unsuccessful.
NEXT: Weight & Measurements.
|
|
|
|
Post by grrraaahhh on Apr 27, 2011 12:06:07 GMT -9
Southwest Yukon/Interior B.C. (continued)Weight (text extract)1. Weights, measurements and pelage1.1. Weights and measurements During the five years of the study, we captured 53 different grizzlies on the study area. The sex ratio was 36 males: 17 females. During the same period, we also captured 31 different black bears (23 males: 8 females) on the area. Table 3 shows the capture success for grizzly bears throughout the study. The total number of grizzlies captured was influenced by two factors. First, certain grizzlies became accustomed to our trapping techniques and were less easily caught. Second, in the last two years, increased captures from helicopters more than offset this reduced success. The measurements for all grizzlies captured during the study are presented in Appendix A (males) and Appendix B (females). Body weights of grizzlies captured are correlated by age for each sex (Table 4). The annual fluctuations will be discussed later. Males were heavier than females of corresponding age in all but cubs and yearlings. Unfortunately, since so few young females were captured during the study, an accurate description of weight patterns during the early years was not possible. The average weight of 40 mature males was 139 kg whereas that of 21 females was 95 kg. The maximum size of each sex shows even greater dimorphism. The largest male weighed 240 kg, nearly twice the weight of the largest female (125 kg). The large boar class can be recognized from size alone when observed free-roaming. It is not likely that many northern interior grizzlies reach a weight greater than the 240 kg recorded in the study. Weights were also obtained of several mature males killed by hunters throughout the Yukon. The five heaviest weighed 218, 203,192,175, and 164 kg. All were killed in the late autumn and should therefore have reflected the heaviest annual weight. All were described as very large by both guide and hunter. Mature females probably never weigh more than 140 kg; the largest killed by hunters weighed 125, 120, 109 and 104 kg. 1.2. Seasonal weight changes The extremes in energy regimes experienced by northern interior grizzly bears during the annual cycle induce major fluctuations in body weight. For at least 6 months, the animals have to survive from energy produced within the body by the utilization offal deposits laid down during the active period. Black bears can increase in weight rapidly when provided with sufficient energy (Cowan et al. 1957; Rausch 1961; Jonkel and Cowan 1971). Jonkel and Cowan (1971) reported significant weight increases during the summer in adult female black bears but not in adult males. Their speculation that the adult males endure the winter denning period more efficiently than the females, and thus have less weight to gain back, did not seem to be the case in grizzly bears in this study. In the late autumn of 1968, we captured four animals (2 adult males; 2 subadult females) shortly before they entered their dens. They were recaptured in the spring of 1969 as they emerged from the winter sleep. Figure 2 shows the astonishing weight changes. The average rate of weight loss for the four animals was 0.20 kg per day per animal for an average period of 220 days. One other adult male was captured in July 1968 (before the period when the animals add fat rapidly) and again in early May 1969. Its projected weight change is also shown in Figure 2. No adult females were measured for the same time span but inasmuch as the four animals lost 30.2, 28.8, 43.2 and 28.0 per cent of their fall weights respectively, it is improbable that the adult female class would lose more and still survive. No mortality was recorded in the adult female class. It is difficult to estimate the rate of weight increase during the period in the autumn when the animals were feeding on soapberries. It is obvious that, given a food source, bears are capable of assimilating energy and increasing in weight rapidly. 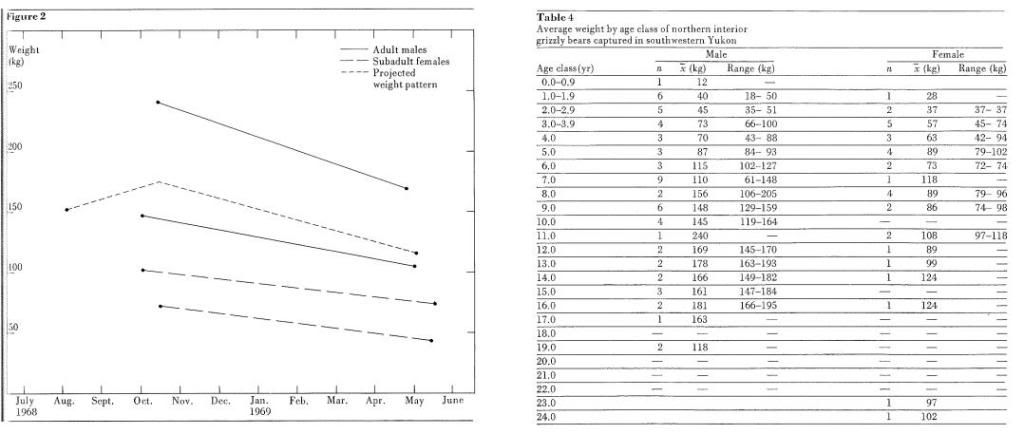 One adult male increased at an average rate of 0.413 kg per day over 126 days. The most rapid increase recorded was of a free-roaming immature female that added weight at 0.635 kg per day over a 16-day period between August 10 and 26. At this time, the bears were feeding exclusively on soapberries. NEXT: Measurements (Skulls)
|
|