|
|
Post by grrraaahhh on Mar 22, 2010 20:39:49 GMT -9
|
|
|
|
Post by grrraaahhh on Mar 25, 2010 6:02:21 GMT -9
|
|
|
|
Post by grrraaahhh on Nov 11, 2010 7:45:57 GMT -9
Family Ursidae
Subfamily Ailuropodinae
+ †Ailurarctos
†Ailurarctos lufengensis
†Ailurarctos yuanmouenensis
+ Ailuropoda (pandas)
†Ailuropoda baconi
†Ailuropoda fovealis
Ailuropoda melanoleuca, giant panda
* Ailuropoda melanoleuca melanoleuca, giant panda
* Ailuropoda melanoleuca qinlingensis, Qinling panda
†Ailuropoda microta
†Ailuropoda wulingshanensis
Subfamily Tremarctinae
+ Tremarctos (Spectacled bears)
Tremarctos ornatus, spectacled bear
†Tremarctos floridanus
+ †Arctodus
†Arctodus simus
†Arctodus pristinus
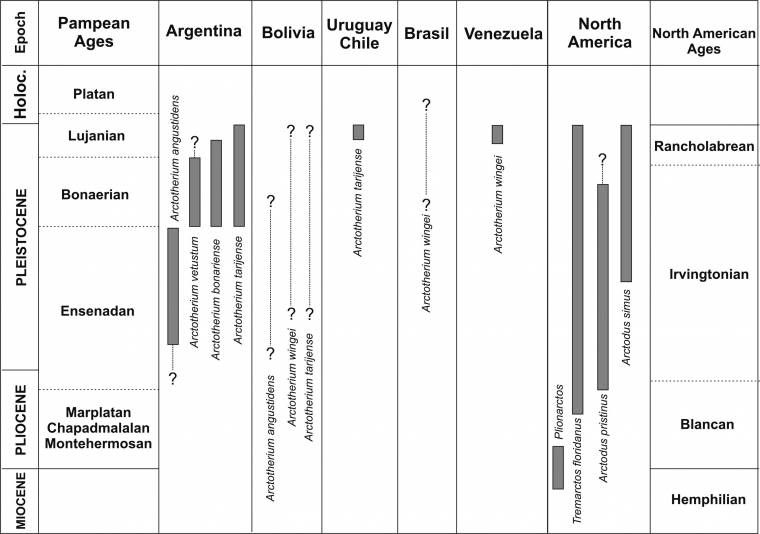
+ †Arctotherium
†Arctotherium angustidens
†Arctotherium bonariense
†Arctotherium brasilense
†Arctotherium latidens
†Arctotherium tarijense
†Arctotherium vetustum
†Arctotherium wingei
+ †Plionarctos
†Plionarctos edensis
†Plionarctos harroldorum
Subfamily Ursinae
+ †Agriotherium
†Agriotherium inexpetans
†Agriotherium schneideri
†Agriotherium sivalensis
+ Helarctos
Helarctos malayanus, Sun bear
* Helarctos malayanus malayanus
* Helarctos malayanus euryspilus, Borneo sun bear
+ †Indarctos
†Indarctos anthraciti
†Indarctos arctoides
†Indarctos atticus
†Indarctos nevadensis
†Indarctos oregonensis
†Indarctos salmontanus
†Indarctos vireti
†Indarctos zdanskyi
+ Melursus
Melursus ursinus, sloth bear
* Melursus ursinus inornatus, Sri Lankan sloth bear
* Melursus ursinus ursinus, Indian sloth bear
+ †Ursavus
†Ursavus brevirhinus
†Ursavus depereti
†Ursavus elmensis
†Ursavus pawniensis
†Ursavus primaevus
+ Ursus
†Ursus abstrusus
Ursus americanus, American black bear
* Ursus americanus altifrontalis, Olympic black bear
* Ursus americanus amblyceps, New Mexico black bear
* Ursus americanus americanus, Eastern black bear
* Ursus americanus californiensis, California black bear
* Ursus americanus carlottae, Haida Gwaii black bear or Queen Charlotte black bear
* Ursus americanus cinnamomum, cinnamon bear
* Ursus americanus emmonsii, Glacier bear
* Ursus americanus eremicus, Mexican black bear
* Ursus americanus floridanus, Florida black bear
* Ursus americanus hamiltoni, Newfoundland black bear
* Ursus americanus kermodei, Kermode bear or spirit bear
* Ursus americanus luteolus, Louisiana black bear
* Ursus americanus machetes, West Mexico black bear
* Ursus americanus perniger, Kenai black bear
* Ursus americanus pugnax, Dall black bear
* Ursus americanus vancouveri, Vancouver Island black bear
# Ursus arctos, brown bear
* Ursus arctos arctos, Eurasian brown bear
* Ursus arctos alascensis
* Ursus arctos beringianus, Kamchatka brown bear or Far Eastern brown bear
* †Ursus arctos californicus, California golden bear
* †Ursus arctos crowtheri, Atlas bear
* †Ursus arctos dalli
* Ursus arctos gobiensis, Gobi bear (very rare)
* Ursus arctos horribilis, grizzly bear
* Ursus arctos isabellinus, Himalayan brown bear or Himalayan red bear
* Ursus arctos lasiotus, Ussuri brown bear or black grizzly
* Ursus arctos middendorffi, Kodiak bear
* †Ursus arctos nelsoni, Mexican grizzly bear
* Ursus arctos piscator, Bergman's bear (extinct)
* Ursus arctos pruinosus, Tibetan blue bear or Tibetan bear or Himalayan blue bear
* Ursus arctos sitkensis
* Ursus arctos syriacus Syrian (brown) bear
†Ursus deningeri
†Ursus etruscus
†Ursus inopinatus, MacFarlane's bear (cryptid; if an authentic species, extinct)
†Ursus minimus
Ursus maritimus, polar bear
* Ursus maritimus maritimus
* †Ursus maritimus tyrannus
†Ursus rossicus
†Ursus sackdillingensis
†Ursus spelaeus, cave bear
Ursus thibetanus, Asian black bear
* Ursus thibetanus formosanus, Formosan black bear
* Ursus thibetanus gedrosianus
* Ursus thibetanus japonicus
* Ursus thibetanus laniger
* Ursus thibetanus mupinensis
* Ursus thibetanus thibetanus
* Ursus thibetanus ussuricus
o †Kolponomos
+ †Kolponomos clallamensis
+ †Kolponomos newportensis
|
|
|
|
Post by grrraaahhh on Nov 11, 2010 7:46:56 GMT -9
|
|
|
|
Post by grrraaahhh on Nov 11, 2010 7:47:33 GMT -9
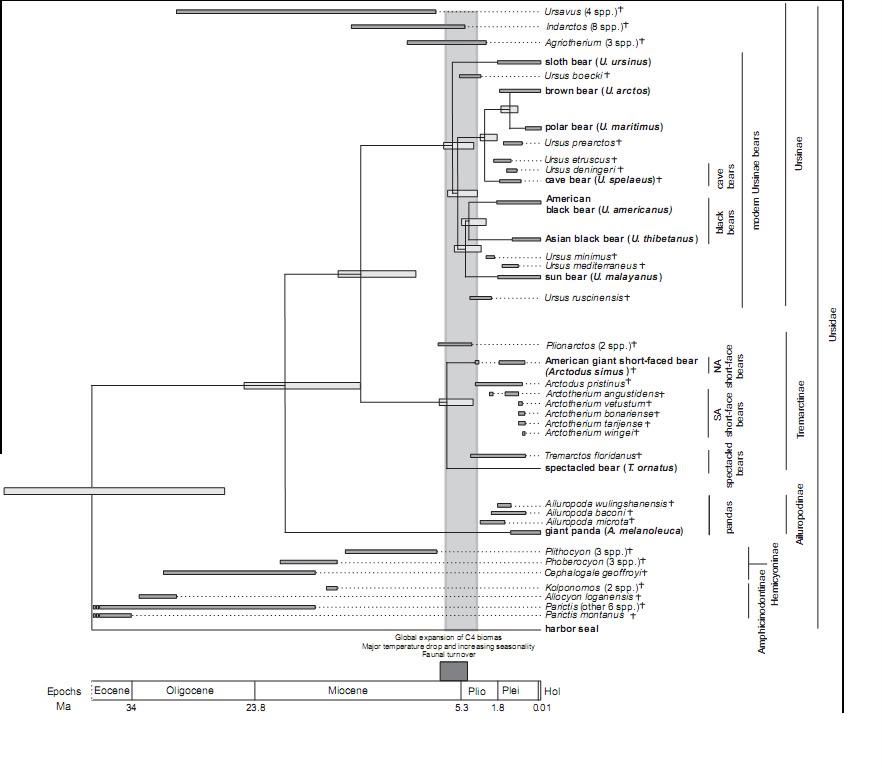
ConclusionSequences from extinct bears represent the third and fourth Pleistocene species for which complete mitochondrial genomes have been sequenced. Moreover, the cave bear specimen demonstrates that mitogenomic studies can be applied to Pleistocene fossils that have not been preserved in permafrost, and therefore have a broad application within ancient DNA research. Molecular dating of the mtDNA divergence times suggests a rapid radiation of bears in both the Old and New Worlds around 5 million years ago, at the Miocene-Pliocene boundary. This coincides with major global changes, such as the Messinian crisis and the first opening of the Bering Strait, and suggests a global influence of such events on species radiations. Mitochondrial genomes reveal an explosive radiation of extinct and extant bears near the Miocene-Pliocene boundarywww.biomedcentral.com/content/pdf/1471-2148-8-220.pdf
|
|
|
|
Post by grrraaahhh on Jan 15, 2011 15:10:13 GMT -9
|
|
|
|
Post by arctodus on Feb 15, 2011 9:17:43 GMT -9
Do you know which is the animal has generated the ursidae family?
thanks.
|
|
|
|
Post by grrraaahhh on Feb 15, 2011 14:21:52 GMT -9
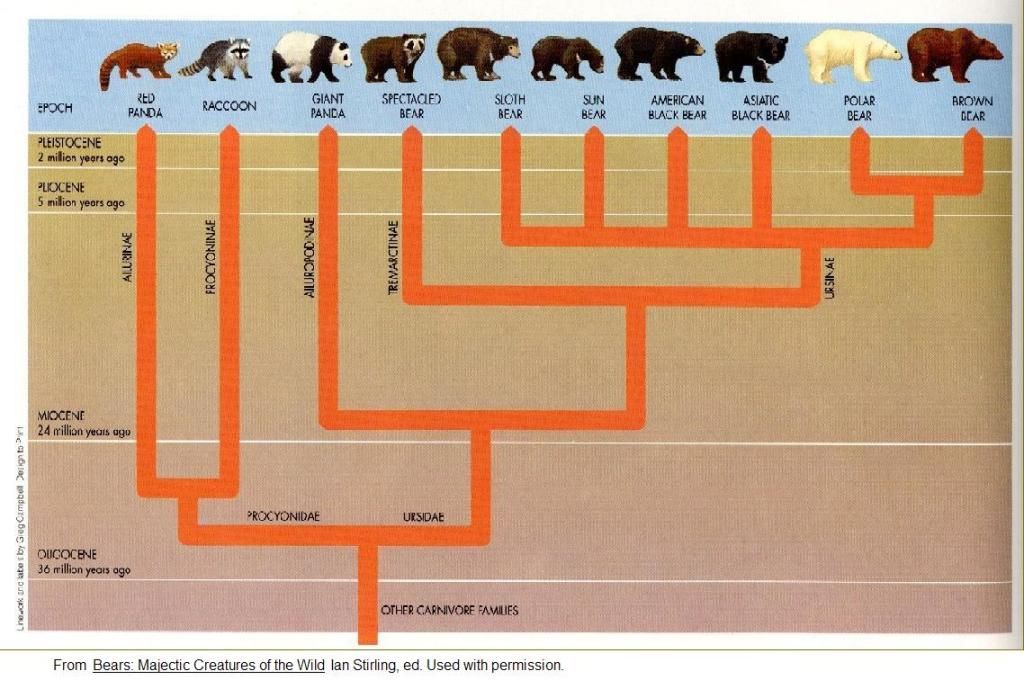
Do you know which is the animal which has generated the ursidae family? thanks. If I understand correctly - the question: Evolutionary Divergence of Ursidae- Origins of this family can be traced to Paractis (very small, skull 7 cm.)
- Paractis first occurred in North America during the Late Eocene, but not found in Eurasia and Africa until Miocene
- However, suggestions leads to speculation that earlier ursids in Asia, during Eocene, gave rise to Paractis in North America, although no fossils of Paractis have been found in East Asia
- This migration may have occurred via a major sea-level lowstand 37 million years ago
- The most primitive ursid was Cephalogale, a raccoon-sized, dog like creature
- Cephalogale first appeared in the Oligocene to early Miocene in Europe
- Cephalogale gave rise to early bears (Ursavus) in Europe
- Ursavus radiated into Asia and gave rise to the first true bears
(Ursus) in Europe or Asia about 5 million years ago
|
|
|
|
Post by arctodus on Feb 15, 2011 20:51:45 GMT -9
You understood well the question, thanks a lot.
|
|
|
|
Post by arctodus on Feb 17, 2011 3:33:17 GMT -9
I'm very interested about paractis, have you got documentation about it?
I would like to know its diet, its mass and look some photos.
Did it live in the trees?
|
|
|
|
Post by grrraaahhh on Feb 17, 2011 22:13:53 GMT -9
No data, just its reference (Allen MS-PPT, reply # 3) which caught my attention outside of the usual Cephalogale & Ursavus/Dawn Bear material.
|
|
|
|
Post by grrraaahhh on Feb 18, 2011 9:51:32 GMT -9
|
|
|
|
Post by grrraaahhh on Jun 28, 2011 18:17:00 GMT -9
Abstract The present distribution and abundance of the ursids is but an ephemeral reflection of an evolutionary path that began with the first identifiable bear, the dawn bear (Ursavus elmensis), 20 million years ago in the early Miocene epoch. Although the dawn bear was only the size of a fox terrier, by the Pleistocene its descendents had evolved into some of the largest terrestrial carnivores the world has known. Most bear species evolved in the northern hemisphere although some dispersed and reached South America, Africa and Southeast Asia. Each species had to cope with ecological changes that affected interspecific competition or the availability of food. Apparently the black bear was sufficiently adapted to have survived largely unchanged from what it was like a million years ago. Numerous species went extinct, leaving only the 8 still present today. Some understanding of the evolutionary pressures that the modern bears have evolved through may help us to understand their behavioral ecology. During the Pleistocene, bears at higher latitudes grew large and ecologically plastic while those closer to the equator remained small and became ecological specialists, as predicted by Geist's (1987) dispersal theory. Adaptations of the teeth of ancestral bear species allowed them to be both herbivores and carnivores. This allowed them to develop large size and broad ecological plasticity. Large body size enabled bears to conserve heat, capture large prey, defend carrion, travel great distances, and, as vegetation increased in the diet, to survive on qualitatively poorer food. Quantity and quality of available food and the degree of sexual dimorphism influenced the size of the home range and the evolution of social behavior in each species. Bears show a great deal of individual variation in behavior and may exploit different subniches as a result of learned behavior. Slight differences in phenotype may also influence exploitation of subniches. Recent literature indicates that some terrestrial bear species are more active predators than previously thought and some evidence suggests a degree of scaling between the size of bears and the size of their prey. Social signalling appears to have been influenced by life in forest habitats but is not well understood. We give a preliminary interpretation of the social organization of the present day bears through the interactive framework of proximate ecological pressures, phylogenetic history, and learning. There are likely few populations of bears anywhere in the world whose behavior has not been significantly influenced by man. This may confound our understanding of their behavior and ecology. Remaining populations of bears may not be able to adapt successfully to the combined effects of human predation, disappearing habitat, and climatic change. Stirling, I. and AE Derocher. 1990. Factors affecting the evolution and behavioral ecology of the modern bears. International Conference on Bear Research and Management 8:189–204. PDF LINK: www.google.com/url?sa=t&source=web&cd=1&ved=0CBYQFjAA&url=http%3A%2F%2Fwww.bearbiology.com%2Ffileadmin%2Ftpl%2FDownloads%2FURSUS%2FVol_8%2FStirling_Derocher_8.pdf&rct=j&q=FACTORS%20AFFECTING%20THE%20EVOLUTION%20AND%20BEHAVIORAL%20ECOLOGY%20OF%20THE%20MODERN%20BEARS1%20&ei=3JcKTv26MYG4sAOV2M2cCQ&usg=AFQjCNErDU6nx690nhn1EsdE4jImfsuP1Q&cad=rja |
|
|
|
Post by grrraaahhh on Oct 27, 2011 4:41:17 GMT -9
One of the landmark/pioneering papers on black bear evolution and ecological adaptation was authored by Stephen Herrero in 1972. Abstract.Certain behavioural, ecological, morphological, and physiological differences between grizzly/brown bears and black bears are related to the different habitats favored by each species. It is suggested that care of black bear cubs and hence reproductive success for black bears is tied to the forest biome, and that the grizzly/brown bear in branching out from the forest onto non-treed areas such as tundras, grasslands and prairie edges, gained rich new food sources, but also became more aggressive than the black bear, a behavioural adaptation to the cub care in this new habitat. Herrero, S., 1972. Aspects of evolution and adaptation in American black bear (Ursus americanus Pallus) and brown and grizzly bears (Ursus arctos Linne.) of North America. Int. Conf. Bear Res. Manage. 2, 221–231. PDF LINK: www.google.com/url?sa=t&rct=j&q=aspects%20of%20evolution%20and%20adaptation%20in.%20american%20black%20bears%20(ursus%20americanus%20pallas)%20and%20brown%20and%20grizzly%20bears%20(u.%20arctos%20linne.)%20of%20north%20america&source=web&cd=1&ved=0CBoQFjAA&url=http%3A%2F%2Fwww.bearbiology.com%2Ffileadmin%2Ftpl%2FDownloads%2FURSUS%2FVol_2%2FHerrero.pdf&ei=c1WpTp_eO4Tv0gGTn93nAw&usg=AFQjCNF5OG7P8hkaFbmo7mF1IFwi2jhkxw&cad=rja |
|
|
|
Post by grrraaahhh on Nov 7, 2011 6:43:38 GMT -9
Abstract I examined the relationship of diets to skull morphology of extant northern bears and used this information to speculate on diets of the recently extinct cave (Ursus spelaeus) and short-faced (Arctodus simus) bears. Analyses relied upon published skull measurements and food habits of Asiatic (U. thibetanus) and American (U. americanus) black bears, polar bears (U. maritimus), various subspecies of brown bears (U. arctos), and the giant panda (Ailuropoda melanoleuca). Principal components analysis showed major trends in skull morphology related to size, crushing force, and snout shape. Giant pandas, short-faced bears, cave bears, and polar bears exhibited extreme features along these gradients. Diets of brown bears in colder, often non-forested environments were distinguished by large volumes of roots, foliage, and vertebrates, while diets of the 2 black bear species and brown bears occupying broadleaf forests contained greater volumes of mast and invertebrates and overlapped considerably. Fractions of fibrous foods in feces (foliage and roots) were strongly related to skull morphology (R2 = 0.97). Based on this relationship, feces of cave and short-faced bears were predicted to consist almost wholly of foliage, roots, or both. I hypothesized that cave bears specialized in root grubbing. In contrast, based upon body proportions and features of the ursid digestivetract, I hypothesized that skull features associated with crashing; force facilitated a carnivorous rather than herbivorous diet for short-faced bears. Mattson, D.J. 1995. Diet and morphology of extant and recently extinct northern bears. Ursus 10:479-496. PDF LINK: www.bearbiology.com/fileadmin/tpl/Downloads/URSUS/Vol_10/MattsonB_Vol_10.pdf |
|