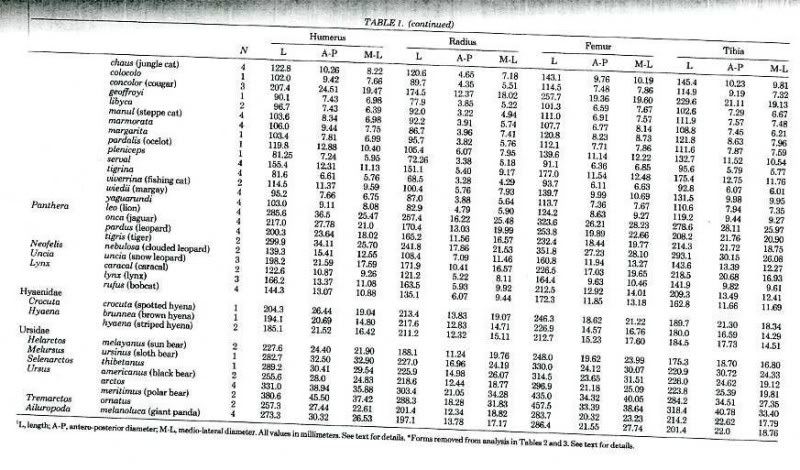
There is large variation in measurements.
Unfortunately, some of them I am posting may be inaccurate.
-Average measure/average measure. Even if both are of the same sample, biases results.
-Measure taken via pixel counting a graph.
-Biased presentation, selection, and focus. Focus on diameter or circumference/length ratios of the humerus of brown bears. I will improve this, but right now my aim is just to post something quickly.
-Measurements may differ between authors
"Scaling Patterns and Ecological Correlates of Postcranial Skeletal Robusticity in Canis and Ursus: Implications for Human Evolution"
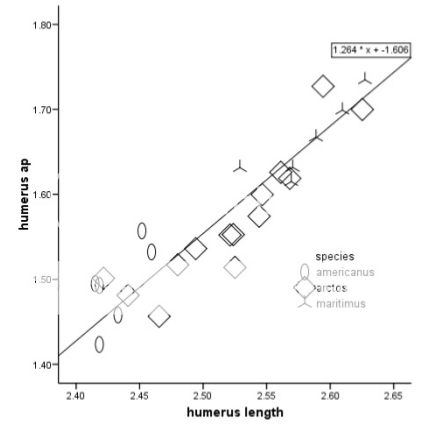

One brown bear's value lies far from that of the others included in this article.
Trying to measure pixels using GIMP:
log(AP diameter): 1.728
AP diameter: 10^1.728= 53.5 mm
log(ML diameter): 1.717
ML diameter= 10^1.717 = 52.1 mm
log(length)= 2.594
Humerus length= 10^2.594 = 393 mm
AP humerus robusticity= 13.6%
ML humerus robusticity= 13.3%
Total humerus robustness= 26.9%
This struck my interest as these values are high. For comparison, the sample of Smilodon fatalis used in "Radiographs Reveal Exceptional Forelimb Strength in the Sabertooth Cat, Smilodon fatalis":
AP%; ML%; Combined%
15.2%; 11.5%; 26.6%
15.0%; 10.8%; 25.8%
15.0%; 9.9%; 24.8%
16.8%; 11.4%; 28.2%
15.0%; 12.2%; 27.2%
14.5%; 10.5%; 25.1%
13.6%; 11.1%; 24.7%
14.0%; 10.9%; 25.0%
15.0%; 11.2%; 26.3%
16.0%; 11.6%; 27.6%
While similar in the combined value, that individual bear was much smaller in the anteroposterior direction, and much greater in mediolateral.
From "Morphofunctional analysis of the postcranium of Amphicyon major (Mammalia, Carnivora, Amphicyonidae) from the Miocene of Sansan (Gers, France) compared to three extant carnivores: Ursus arctos, Panthera leo, and Canis lupus":
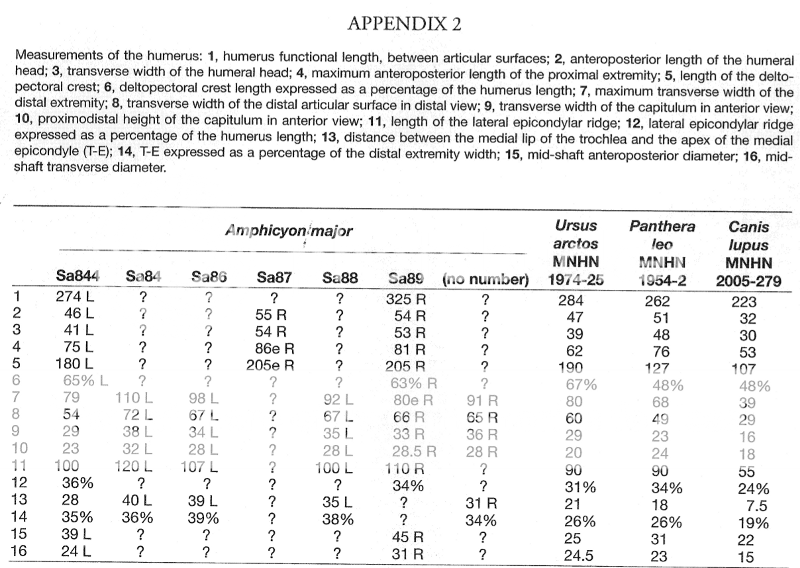
Variation in diameter/length ratios is interesting. Compare AP/length and MP length of 8.8% and 8.6% respectively with the maximum of the bear from the above article.
High variation, probably due to high variation in body mass relative to leg length in bears.
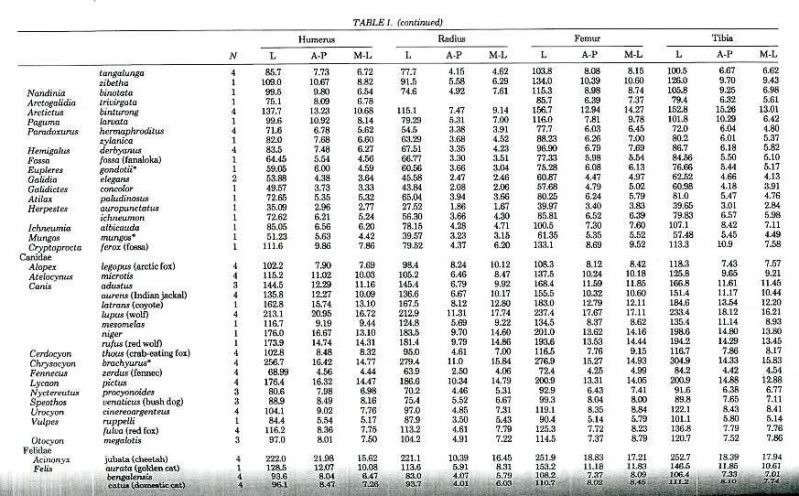
"Differential Scaling of Limb Bones in Terrestrial Carnivores and Other Mammalia" by Biewner includes average values for a large sample of Carnivora. I will add values for a large variety of ungulates within the next week.
The article "What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae)?" by Per Christiansen also includes graphs of fore and hindlimb least circumference and body length.
Here is the graph, and
here is the key.
From using GIMP to pixel count, these are the values I got for brown bears and the "slender legged" Arctodus simus. If there is interest in compiling info in a table more species could be added.
Humerus 1:
785,211
Roughly 145 mm least circumference, and 447 mm long.
Least circumference/length= 0.324
Humerus 2:
806, 207
Humeral least circumference of about 146 mm, and about 457 mm long.
Least circumference/length= 0.319
Humerus 3:
890,177.5
Least circumference of about 154 mm, and about 497 mm long.
Least circumference/length= 0.311
Humerus 4:
1086, 50
Least circumference of about 190 mm, and about 590 mm long.
Least circumference/length= 0.322
Non-Kodiak brown bears:
Humerus 1:
337, 447
Least circumference: 77.3
Length: 234 mm
Least circumference/length= 0.330
Humerus 2:
375, 461
Least circumference: 73.3
Length: 252 mm
Least circumference/length= 0.290
Humeri 1 and 2 belong to the two smallest brown bears. Weight estimates based on their humeri lengths. least circumferences, and average of both respectively are:
Humerus 1: 53, 89, 71 kg
Humerus 2: 70, 77, 73 kg
Humerus 3:
385.5, 415.5
Least circumference: 86.4
Length: 257 mm
Least circumference/length= 0.335
Humerus 4:
412, 465
Least circumference: 72.1
Length: 270 mm
Least circumference/length= 0.267
Humerus 5:
413, 433
Least circumference: 81.3
Length: 270 mm
Least circumference/length= 0.301
Humerus 6:
423, 415.5
Least circumference: 86.4
Length: 275 mm
Least circumference/length= 0.314
Humerus 7:
463, 419
Least circumference: 85.3
Length: 294 mm
Least circumference/length= 0.290
Humerus 8:
538, 307
Least circumference: 117
Length: 330 mm
Least circumference/length= 0.356
Weight estimates based on humerus length, least circumference, and the average respectively for the two largest regular brown bears included:
Humerus 7: 123, 115, 119 kg
Humerus 8: 187, 268, 228 kg
Kodiak bears:
Humerus 1:
580, 279
Least circumference: 125
Length: 350 mm
Least circumference/length= 0.358
Humerus 2:
731, 191
Least circumference: 151
Length: 422 mm
Least circumference/length= 0.357
Weights of the two Kodiak bears based on humerus lengths, least circumference, and average of the two respectively:
Humerus 1: 233, 320, 276 kg
Humerus 2: 463, 518, 490 kg
The two Kodiak bears and the largest of the brown bears (but still only modest) had large least circumference to length ratios.
Arctodus simus specimens were far from slender.
For reference, "Osteology and ecology of Megantereon cultridens SE311 (Mammalia; Felidae; Machairodontinae), a sabrecat from the Late Pliocene – Early Pleistocene of Senéze, France" has average values for felines. Remember though that felines have humeri far more oval in cross section than those of bears however.
As in other derived sabrecats, the humerus of
Megantereon is very similar to those of extant big
cats, albeit with exaggerated robustness overall.
The ratio of humerus least circumference of the dia-
physis relative to articular length in Megantereon
(0.356; Fig. 15A) is distinctly higher than in Panthera
leo (0.318 ± 0.005; P < 0.001), P. onca (0.322 ± 0.008;
P < 0.001), P. pardus (0.290 ± 0.005; P < 0.001),
P. tigris (0.303 ± 0.005; P < 0.001), Neofelis nebulosa
(0.298 ± 0.010; P = 0.009) and Puma concolor
(0.291 ± 0.009; P < 0.001), and even than in Smilodon
gracilis (0.330 ± 0.011; P = 0.047) and S. fatalis
(0.337 ± 0.008; P = 0.027). Only the very robust
S. populator has a higher circumference/length ratio
than Megantereon (0.401 ± 0.011; P = 0.003).Note again the very high variation from brown bear to brown bear- the brown bear's standard deviation, at ± 0.02896, was much higher than those of any of these members of Felidae.
Smilodon populator had an extremely robust humerus.
On the reliability of my "pixel counting": questionable. I got 385 kg instead of 389 kg from the table in "Morphometric characteristics of brown bears on the central Alaska Peninsula". Mistakes are amplified when converting from log graphs.
Per Christiansen used the same sample of bones in his article "Scaling of mammalian long bones: small and large
mammals compared".
Average least circumference value of the two Kodiak bears: 138.5 mm, vs 138 I got from the graph.
Average humerus length: 386.0 vs 386. Cool!
Actual average vs one I got for:
Brown bear humeral least circumference: 85.6 vs 84.8875
Length: 272.8 vs 272.75
Not perfect, but IMO it is reasonable.
Small sample from supporting info of "A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods":
Common name Specimen Body mass H length H circumference F length F least circumference
Black bear ROM 71435 204000 296 101.5 302 96.5
Grizzly bear ROM 35699 435500 400.5 146.25 445.5 126.25
Polar bear AHR1985 447695 158 135
Measurements are g and mm.
The grizzly included in the above article's humerus least circumference/length ratio was 0.365.


